

Uncoupling and turnover in a Cl-/H+ exchange transporter
- PMID: 17389248
- PMCID: PMC2151619
- DOI: 10.1085/jgp.200709756
Uncoupling and turnover in a Cl-/H+ exchange transporter
Abstract
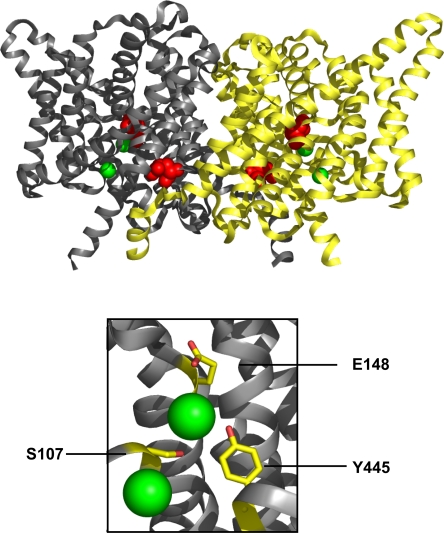
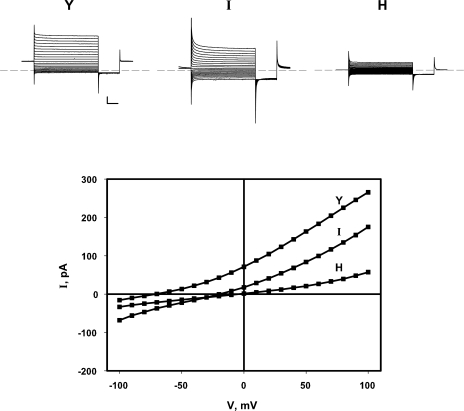
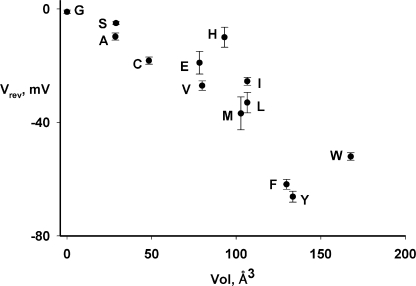
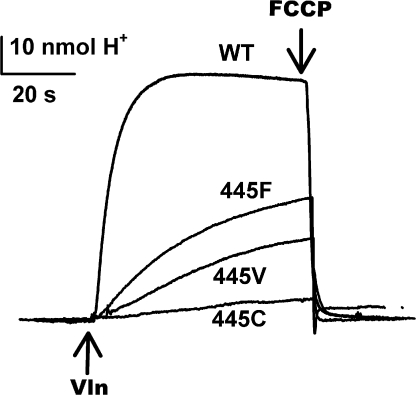
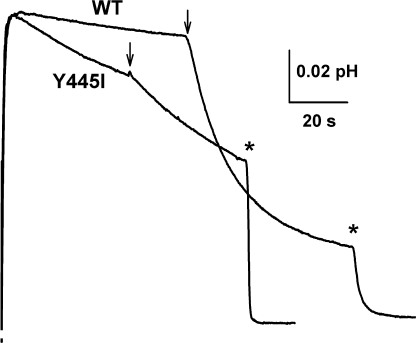
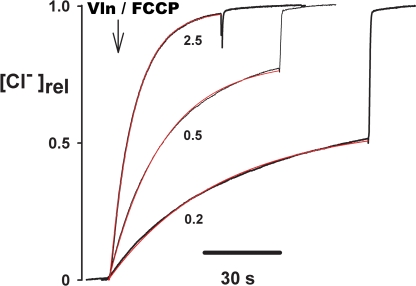
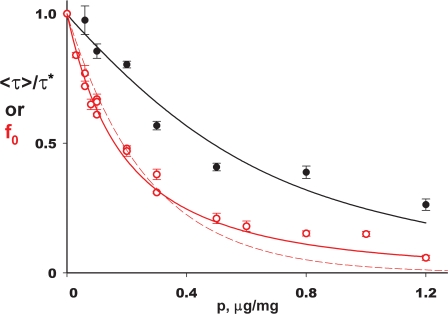
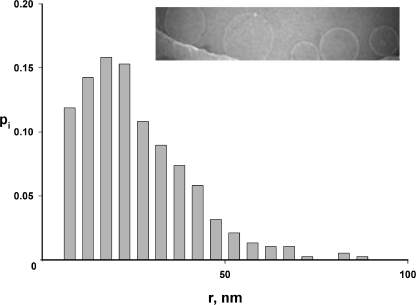
The CLC-family protein CLC-ec1, a bacterial homologue of known structure, stoichiometrically exchanges two Cl(-) for one H(+) via an unknown membrane transport mechanism. This study examines mutations at a conserved tyrosine residue, Y445, that directly coordinates a Cl(-) ion located near the center of the membrane. Mutations at this position lead to "uncoupling," such that the H(+)/Cl(-) transport ratio decreases roughly with the volume of the substituted side chain. The uncoupled proteins are still able to pump protons uphill when driven by a Cl(-) gradient, but the extent and rate of this H(+) pumping is weaker in the more uncoupled variants. Uncoupling is accompanied by conductive Cl(-) transport that is not linked to counter-movement of H(+), i.e., a "leak." The unitary Cl(-) transport rate, measured in reconstituted liposomes by both a conventional initial-velocity method and a novel Poisson dilution approach, is approximately 4,000 s(-1) for wild-type protein, and the uncoupled mutants transport Cl(-) at similar rates.
Figures

References
-
- Accardi, A., S. Lobet, C. Williams, C. Miller, and R. Dutzler. 2006. Synergism between halide binding and proton transport CLC-type exchanger. J. Mol. Biol. 362:691–699. - PubMed
-
- Accardi, A., and C. Miller. 2004. Secondary active transport mediated by a prokaryotic homologue of ClC Cl− channels. Nature. 427:803–807. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
