

Trans-splicing of the Ll.LtrB group II intron in Lactococcus lactis
- PMID: 17389638
- PMCID: PMC1874635
- DOI: 10.1093/nar/gkl1146
Trans-splicing of the Ll.LtrB group II intron in Lactococcus lactis
Abstract
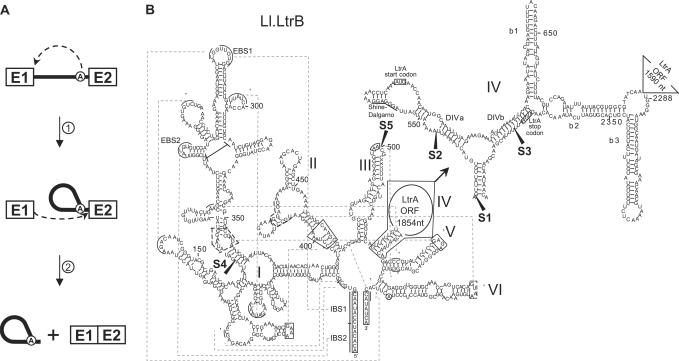
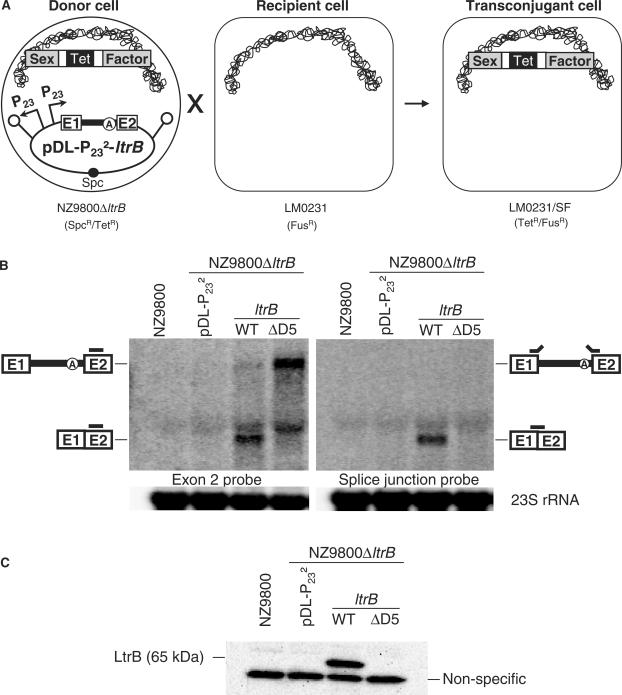
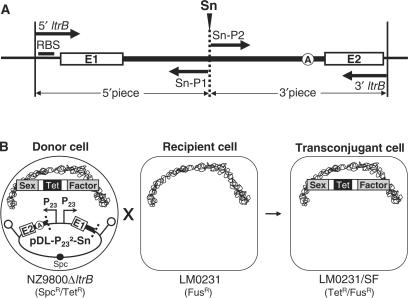
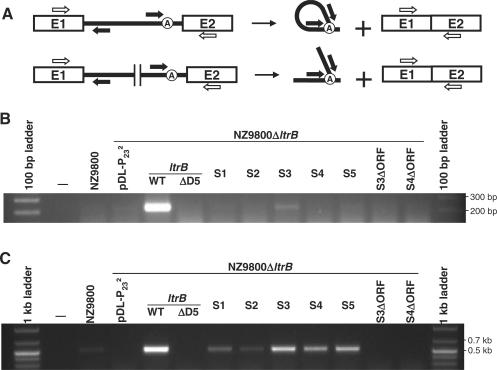
The Ll.LtrB intron from the Gram-positive bacterium Lactococcus lactis is one of the most studied bacterial group II introns. Ll.LtrB interrupts the relaxase gene of three L. lactis conjugative elements. The relaxase enzyme recognizes the origin of transfer (oriT ) and initiates the intercellular transfer of its conjugative element. The splicing efficiency of Ll.LtrB from the relaxase transcript thus controls the conjugation level of its host element. Here, we used the level of sex factor conjugation as a read-out for Ll.LtrB splicing efficiency. Using this highly sensitive splicing/conjugation assay (10(7)-fold detection range), we demonstrate that Ll.LtrB can trans-splice in L. lactis when fragmented at various positions such as: three different locations within domain IV, within domain I and within domain III. We also demonstrate that the intron-encoded protein, LtrA, is absolutely required for Ll.LtrB trans-splicing. Characteristic Y-branched trans-spliced introns and ligated exons are detected by RT-PCR from total RNA extracts of cells harbouring fragmented Ll.LtrB. The splicing/conjugation assay we developed constitutes the first model system to study group II intron trans-splicing in vivo. Although only previously observed in bacterial-derived organelles, we demonstrate that assembly and trans-splicing of a fragmented group II intron can take place efficiently in bacterial cells.
Figures

Similar articles
-
Conjugative transfer of the Lactococcus lactis chromosomal sex factor promotes dissemination of the Ll.LtrB group II intron.J Bacteriol. 2005 Feb;187(3):930-9. doi: 10.1128/JB.187.3.930-939.2005. J Bacteriol. 2005. PMID: 15659671 Free PMC article.
-
Restriction for gene insertion within the Lactococcus lactis Ll.LtrB group II intron.RNA. 2006 Nov;12(11):1980-92. doi: 10.1261/rna.193306. Epub 2006 Sep 14. RNA. 2006. PMID: 16973892 Free PMC article.
-
Group II introns and expression of conjugative transfer functions in lactic acid bacteria.Antonie Van Leeuwenhoek. 1999 Jul-Nov;76(1-4):77-88. Antonie Van Leeuwenhoek. 1999. PMID: 10532373 Review.
-
Conjugation as a Highly Sensitive Assay to Study Group II Intron Splicing In Vivo.Methods Mol Biol. 2021;2167:3-11. doi: 10.1007/978-1-0716-0716-9_1. Methods Mol Biol. 2021. PMID: 32712911
-
Bacterial group II introns and their association with mobile genetic elements.Front Biosci. 2002 Aug 1;7:d1843-56. doi: 10.2741/klein1. Front Biosci. 2002. PMID: 12133822 Review.
Cited by
-
Bacterial group II introns generate genetic diversity by circularization and trans-splicing from a population of intron-invaded mRNAs.PLoS Genet. 2018 Nov 21;14(11):e1007792. doi: 10.1371/journal.pgen.1007792. eCollection 2018 Nov. PLoS Genet. 2018. PMID: 30462638 Free PMC article.
-
Therapeutic applications of trans-splicing.Br Med Bull. 2020 Dec 15;136(1):4-20. doi: 10.1093/bmb/ldaa028. Br Med Bull. 2020. PMID: 33010155 Free PMC article.
-
Contribution of base-pairing interactions between group II intron fragments during trans-splicing in vivo.RNA. 2011 Dec;17(12):2212-21. doi: 10.1261/rna.028886.111. Epub 2011 Oct 27. RNA. 2011. PMID: 22033330 Free PMC article.
-
Trans-splicing versatility of the Ll.LtrB group II intron.RNA. 2008 Sep;14(9):1782-90. doi: 10.1261/rna.1083508. Epub 2008 Jul 22. RNA. 2008. PMID: 18648072 Free PMC article.
-
Alternative splicing of a group II intron in a surface layer protein gene in Clostridium tetani.Nucleic Acids Res. 2014 Feb;42(3):1959-69. doi: 10.1093/nar/gkt1053. Epub 2013 Nov 8. Nucleic Acids Res. 2014. PMID: 24214997 Free PMC article.
References
-
- Michel F, Ferat JL. Structure and activities of group II introns. Annu. Rev. Biochem. 1995;64:435–461. - PubMed
-
- Lambowitz AM, Zimmerly S. Mobile group II introns. Annu. Rev. Genet. 2004;38:1–35. - PubMed
-
- Lehmann K, Schmidt U. Group II introns: structure and catalytic versatility of large natural ribozymes. Crit Rev. Biochem. Mol. Biol. 2003;38:249–303. - PubMed
-
- Sharp PA. Five easy pieces. Science. 1991;254:663. - PubMed
