

Dynamic binding of Ku80, Ku70 and NF90 to the IL-2 promoter in vivo in activated T-cells
- PMID: 17389650
- PMCID: PMC1874627
- DOI: 10.1093/nar/gkm117
Dynamic binding of Ku80, Ku70 and NF90 to the IL-2 promoter in vivo in activated T-cells
Abstract
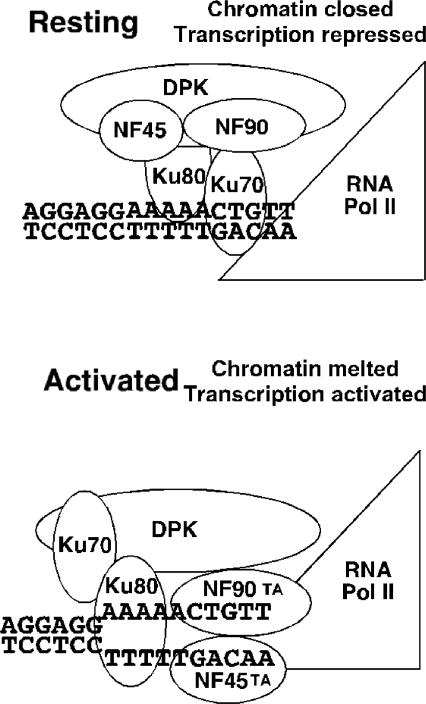
IL-2 gene expression in activated T-cells is initiated by chromatin remodeling at the IL-2 proximal promoter and conversion of a transcriptional repressor into a potent transcriptional activator. A purine-box regulator complex was purified from activated Jurkat T-cell nuclei based on sequence-specific DNA binding to the antigen receptor response element (ARRE)/nuclear factor of activated T-cells (NF-AT) target DNA sequence in the proximal IL-2 promoter. ARRE DNA-binding subunits were identified as NF90, NF45 and systemic lupus erythematosis autoantigens, Ku80 and Ku70. Monoclonal antibodies to Ku80, Ku70 and NF90 specifically inhibit constitutive and inducible ARRE DNA-binding activity in Jurkat T-cells. Ku80, Ku70 and NF90 bind specifically to the IL-2 gene promoter in vivo, as demonstrated by chromatin immunoprecipitation. Activation of Jurkat T-cells and mouse primary spleen cells induces binding of Ku80 and NF90 to the IL-2 promoter in vivo, and decreases binding of Ku70 to the IL-2 promoter in vivo, and these dynamic changes are inhibited by immunosuppressants cyclosporin A and triptolide. Dynamic changes in binding of Ku80, Ku70 and NF90 to the IL-2 proximal promoter in vivo correlate with chromatin remodeling and transcriptional initiation in activated T-cells.
Figures
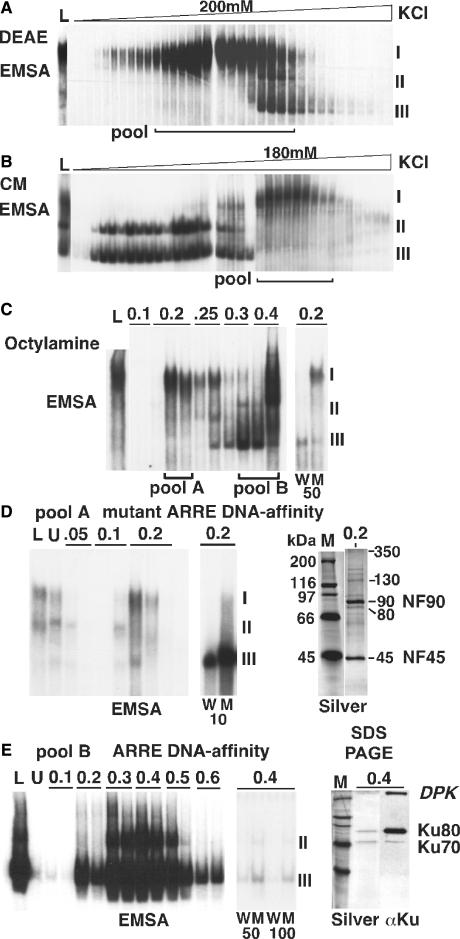
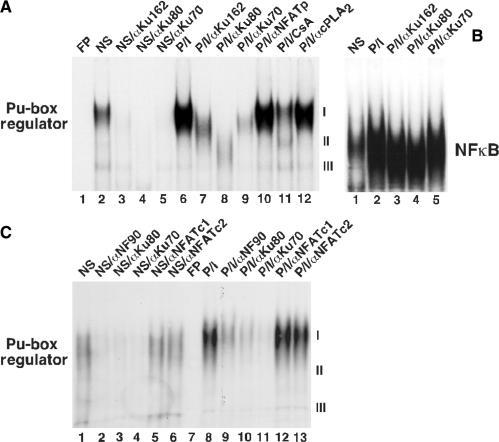
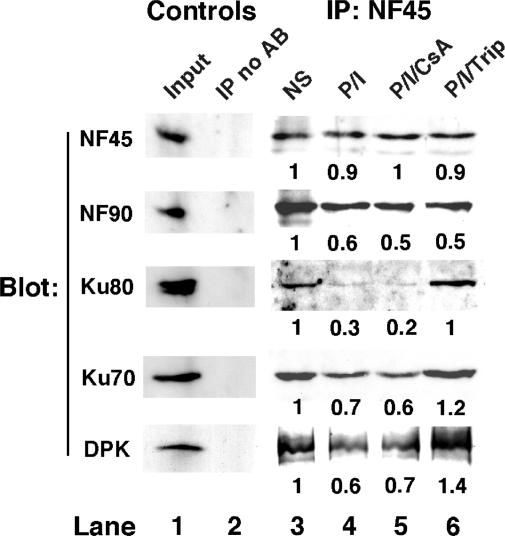
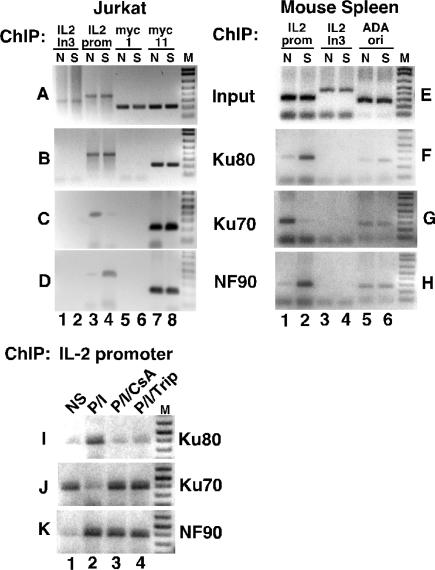
References
-
- June CH, Ledbetter JA, Lindsten T, Thompson CB. Evidence for the involvement of three distinct signals in the induction of IL-2 gene expression in human T lymphocytes. J. Immunol. 1989;143:153–161. - PubMed
-
- Mouzaki A, Rungger D. Properties of transcription factors regulating interleukin-2 gene transcription through the NFAT binding site in untreated or drug-treated naive and memory T-helper cells. Blood. 1994;84:2612–2621. - PubMed
-
- Shaw JP, Utz PJ, Durand DB, Toole JJ, Emmel EA, Crabtree GR. Identification of a putative regulator of early T cell activation genes. Science. 1988;241:202–205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
