

Regulation of protein kinase Mzeta synthesis by multiple kinases in long-term potentiation
- PMID: 17392460
- PMCID: PMC6672124
- DOI: 10.1523/JNEUROSCI.5612-06.2007
Regulation of protein kinase Mzeta synthesis by multiple kinases in long-term potentiation
Abstract
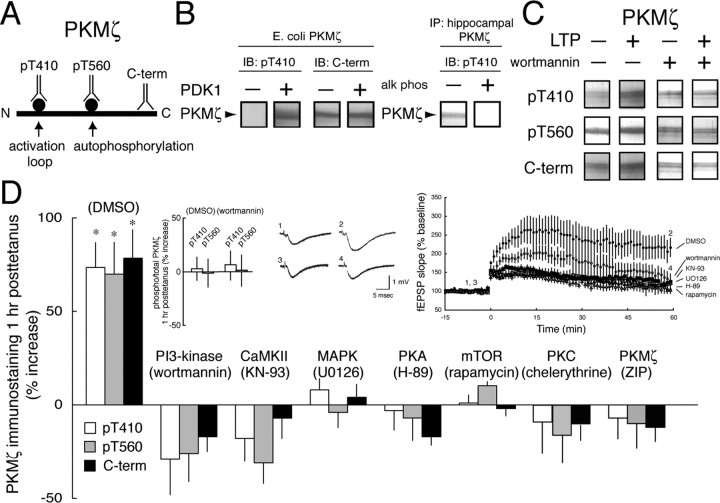
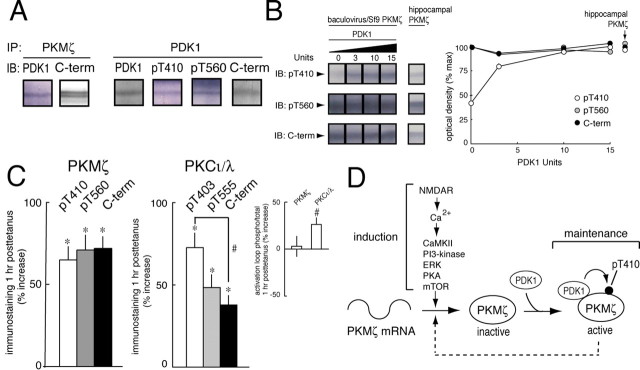
The persistent activity of protein kinase Mzeta (PKMzeta) maintains synaptic long-term potentiation (LTP) and spatial memory, but the interactions between PKMzeta and the other protein kinases implicated in synaptic plasticity are unknown. During LTP, PKMzeta is rapidly synthesized from a PKMzeta mRNA that encodes a protein kinase Czeta (PKCzeta) catalytic domain without a regulatory domain; thus, second messengers that activate full-length PKC isoforms are not required to stimulate PKMzeta. Like other PKCs, however, PKMzeta must be phosphorylated on its activation loop by phosphoinositide-dependent protein kinase-1 (PDK1) for optimal catalytic activity. Thus, two sequential steps are required for the persistent increased PKMzeta activity that maintains LTP: de novo synthesis of PKMzeta and phosphorylation of its activation loop. Here, using a panel of antisera to phosphorylated and nonphosphorylated sites on PKMzeta, we show that PI3-kinase (phosphoinositide 3-kinase), CaMKII (Ca2+/calmodulin-dependent protein kinase II), MAPK (mitogen-activated protein kinase), PKA (protein kinase A), mTOR (mammalian target of rapamycin), all important for LTP induction, as well as preexisting PKMzeta, regulate the new synthesis of PKMzeta during LTP. In contrast, PDK1 forms a complex with PKMzeta and maintains maximal phosphorylation of its activation loop. Thus, the two steps of PKMzeta formation serve separate functions in LTP: the initial regulated synthesis of PKMzeta is the site of convergence and integration for multiple kinases of induction, whereas the constitutive phosphorylation of PKMzeta by PDK1 initiates the persistent autonomous activity that sustains maintenance.
Figures
References
-
- Balendran A, Biondi RM, Cheung PC, Casamayor A, Deak M, Alessi DR. A 3-phosphoinositide-dependent protein kinase-1 (PDK1) docking site is required for the phosphorylation of protein kinase Cζ (PKCζ) and PKC-related kinase 2 by PDK1. J Biol Chem. 2000a;275:20806–20813. - PubMed
-
- Balendran A, Hare GR, Kieloch A, Williams MR, Alessi DR. Further evidence that 3-phosphoinositide-dependent protein kinase-1 (PDK1) is required for the stability and phosphorylation of protein kinase C (PKC) isoforms. FEBS Lett. 2000b;484:217–223. - PubMed
-
- Biondi RM. Phosphoinositide-dependent protein kinase 1, a sensor of protein conformation. Trends Biochem Sci. 2004;29:136–142. - PubMed
-
- Bliss TV, Collingridge GL. A synaptic model of memory: long-term potentiation in the hippocampus. Nature. 1993;361:31–39. - PubMed
-
- Bliss TV, Collingridge GL, Laroche S. Neuroscience. ZAP and ZIP, a story to forget. Science. 2006;313:1058–1059. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous