

Gap junctions contain different amounts of cholesterol which undergo unique sequestering processes during fiber cell differentiation in the embryonic chicken lens
- PMID: 17392685
- PMCID: PMC2642917
Gap junctions contain different amounts of cholesterol which undergo unique sequestering processes during fiber cell differentiation in the embryonic chicken lens
Abstract
Purpose: To determine the possible changes in the distribution of cholesterol in gap junction plaques during fiber cell differentiation and maturation in the embryonic chicken lens. The possible mechanism by which cholesterol is removed from gap junction plaques is also investigated.
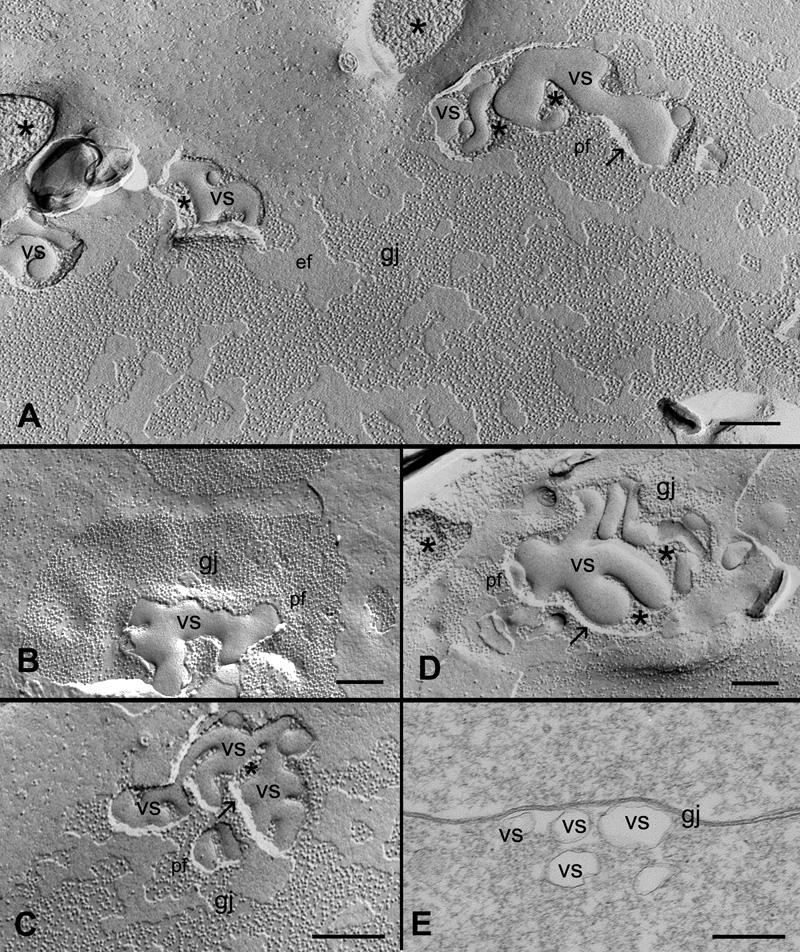
Methods: Filipin cytochemistry in conjunction with freeze-fracture TEM was used to visualize cholesterol, as represented by filipin-cholesterol complexes (FCCs) in gap junction plaques. Quantitative analysis on the heterogeneous distribution of cholesterol in gap junction plaques was conducted from outer and inner cortical regions. A novel technique combining filipin cytochemistry with freeze-fracture replica immunogold labeling (FRIL) was used to label Cx45.6 and Cx56 antibodies in cholesterol-containing gap junctions. Filipin cytochemistry and freeze-fracture TEM and thin-section TEM were used to examine the appearance and nature of the cholesterol-containing vesicular structures associated with gap junction plaques.
Results: Chicken lens fibers contain cholesterol-rich, cholesterol-intermediate and cholesterol-free gap junction populations in both outer and inner cortical regions. Filipin cytochemistry and FRIL studies confirmed that cholesterol-containing junctions were gap junctions. Quantitative analysis showed that approximately 86% of gap junctions in the outer cortical zone were cholesterol-rich gap junctions, whereas approximately 81% of gap junctions in the inner cortical zone were cholesterol-free gap junctions. A number of pleiomorphic cholesterol-rich vesicles of varying sizes were often observed in the gap junction plaques. They appear to be involved in the removal of cholesterol from gap junction plaques through endocytosis.
Conclusions: Gap junctions in the young fibers are enriched with cholesterol because they are assembled in the unique cholesterol-rich cell membranes in the lens. A majority of cholesterol-rich gap junctions in the outer young fibers are transformed into cholesterol-free ones in the inner mature fibers during fiber cell maturation. A distinct endocytotic process appears to be involved in removing cholesterol from the cholesterol-containing gap junctions, and it may play a major role in the transformation of cholesterol-rich gap junctions into cholesterol-free ones during fiber cell maturation.
Figures








Similar articles
-
Gap junction remodeling associated with cholesterol redistribution during fiber cell maturation in the adult chicken lens.Mol Vis. 2009 Aug 4;15:1492-508. Mol Vis. 2009. PMID: 19657477 Free PMC article.
-
Gap junctions are selectively associated with interlocking ball-and-sockets but not protrusions in the lens.Mol Vis. 2010 Nov 9;16:2328-41. Mol Vis. 2010. PMID: 21139982 Free PMC article.
-
Regional changes of AQP0-dependent square array junction and gap junction associated with cortical cataract formation in the Emory mutant mouse.Exp Eye Res. 2014 Oct;127:132-42. doi: 10.1016/j.exer.2014.07.020. Epub 2014 Jul 31. Exp Eye Res. 2014. PMID: 25088353 Free PMC article.
-
Molecular mechanisms regulating formation, trafficking and processing of annular gap junctions.BMC Cell Biol. 2016 May 24;17 Suppl 1(Suppl 1):22. doi: 10.1186/s12860-016-0087-7. BMC Cell Biol. 2016. PMID: 27230503 Free PMC article. Review.
-
Ultrastructure, histological distribution, and freeze-fracture immunocytochemistry of gap junctions in rat brain and spinal cord.Cell Biol Int. 1998 Nov;22(11-12):731-49. doi: 10.1006/cbir.1998.0392. Cell Biol Int. 1998. PMID: 10873288 Review.
Cited by
-
Quantification of Age-Related Changes in the Lateral Organization of the Lipid Portion of the Intact Membranes Isolated from the Left and Right Eye Lenses of the Same Human Donor.Membranes (Basel). 2023 Feb 3;13(2):189. doi: 10.3390/membranes13020189. Membranes (Basel). 2023. PMID: 36837692 Free PMC article.
-
Connexins: a myriad of functions extending beyond assembly of gap junction channels.Cell Commun Signal. 2009 Mar 12;7:4. doi: 10.1186/1478-811X-7-4. Cell Commun Signal. 2009. PMID: 19284610 Free PMC article.
-
Cholesterol and cholesterol bilayer domains inhibit binding of alpha-crystallin to the membranes made of the major phospholipids of eye lens fiber cell plasma membranes.Exp Eye Res. 2021 May;206:108544. doi: 10.1016/j.exer.2021.108544. Epub 2021 Mar 17. Exp Eye Res. 2021. PMID: 33744256 Free PMC article.
-
Gap junction remodeling associated with cholesterol redistribution during fiber cell maturation in the adult chicken lens.Mol Vis. 2009 Aug 4;15:1492-508. Mol Vis. 2009. PMID: 19657477 Free PMC article.
-
Connexin channels and phospholipids: association and modulation.BMC Biol. 2009 Aug 17;7:52. doi: 10.1186/1741-7007-7-52. BMC Biol. 2009. PMID: 19686581 Free PMC article.
References
-
- Cenedella RJ. Sterol synthesis by the ocular lens of the rat during postnatal development. J Lipid Res. 1982;23:619–26. - PubMed
-
- Cenedella RJ. Regional distribution of sterol and fatty acid synthesis in the ocular lens. Exp Eye Res. 1984;38:95–9. - PubMed
-
- Borchman D, Cenedella RJ, Lamba OP. Role of cholesterol in the structural order of lens membrane lipids. Exp Eye Res. 1996;62:191–7. - PubMed
-
- Zelenka PS. Lens lipids. Curr Eye Res. 1984;3:1337–59. - PubMed
-
- Li LK, So L, Spector A. Membrane cholesterol and phospholipid in consecutive concentric sections of human lenses. J Lipid Res. 1985;26:600–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous