

Imaging and manipulating phosphoinositides in living cells
- PMID: 17395624
- PMCID: PMC2075240
- DOI: 10.1113/jphysiol.2007.132795
Imaging and manipulating phosphoinositides in living cells
Abstract
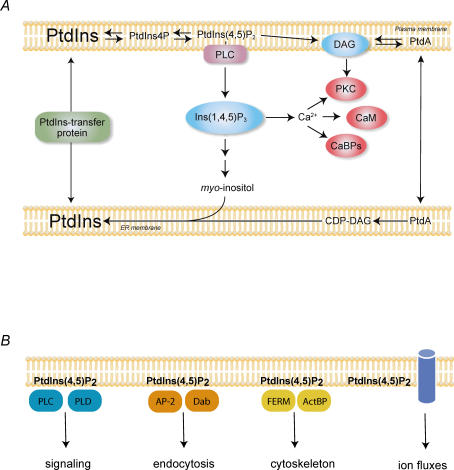
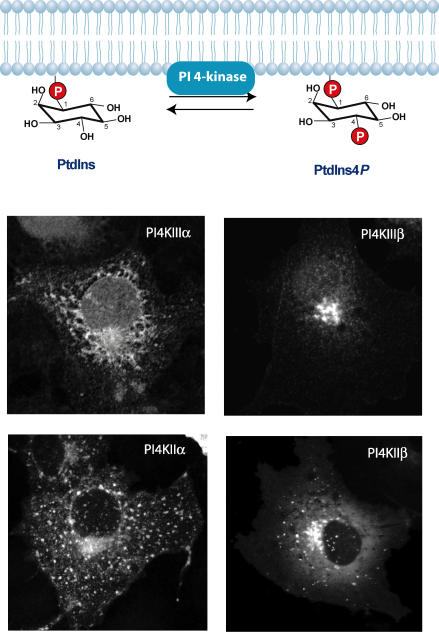
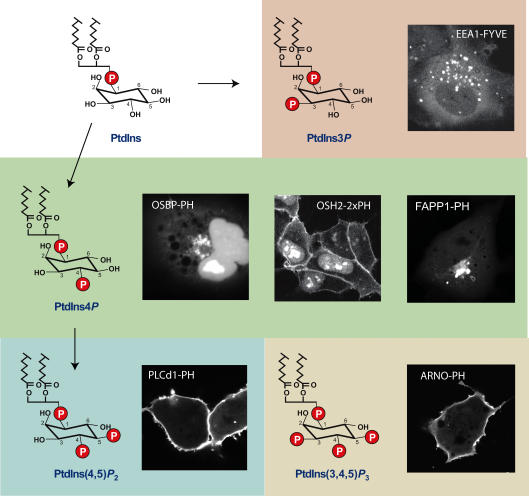
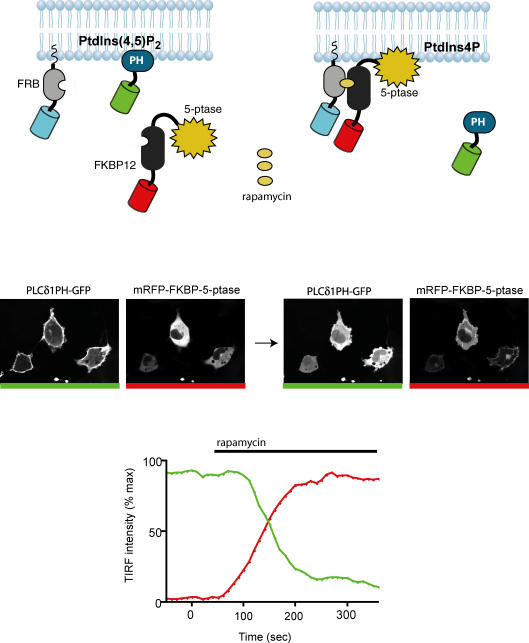
Phosphoinositides are minor phospholipid constituents of virtually every biological membrane yet they play fundamental roles in controlling membrane-bound signalling events. Phosphoinositides are produced from phosphatidylinositol (PtdIns) by phosphorylation of one or more of three positions (3, 4 and 5) of the inositol headgroup located at the membrane cytoplasmic interface by distinct families of inositol lipid kinases. Intriguingly, many of the kinase reactions are catalysed by more than one form of the kinases even in simple organisms and these enzymes often assume non-redundant functions. A similar diversity is seen with inositide phosphatases, the enzymes that dephosphorylate phosphoinositides with a certain degree of specificity and the impairments of which are often linked to human diseases. This degree of multiplicity at the enzyme level together with the universal roles of these lipids in cell regulation assumes that inositol lipids are spatially and functionally restricted in specific membrane compartments. Studying the compartmentalized roles of these lipids at the cellular level represents a major methodological challenge. Over the last 10 years significant progress has been made in creating reagents that can monitor inositol lipid changes in live cells with fluorescence or confocal microscopy. New methods are also being developed to manipulate these lipids in specific membrane compartments in a regulated fashion. This article recalls some historical aspects of inositide research and describes the new methodological advances highlighting their great potential as well as the problems one can encounter with their use.
Figures
References
-
- Astle MV, Seaton G, Davies EM, Fedele CG, Rahman P, Arsala L, Mitchell CA. Regulation of phosphoinositide signaling by the inositol polyphosphate 5-phosphatases. IUBMB Life. 2006;58:451–456. - PubMed
-
- Attree O, Olivos IM, Okabe I, Bauiley LC, Nelson DL, Lewis RA, Mcinnes RR, Nussbaum RL. The Lowe's oculocerebrorenal syndrome gene encodes a protein highly homologous to inositol polyphosphate-5-phosphatase. Nature. 1992;358:239–242. - PubMed
-
- Balla A, Balla T. Phosphatidylinositol 4-kinases; old enzymes with emerging functions. Trends Cell Biol. 2006;16:351–361. - PubMed
-
- Ballou CE, Lee YC. The structure of a myoinositol mannoside from Myobacterium tuberculosis glycolipid. Biochemistry. 1964;3:682–685. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous