

Structural basis for enzymatic excision of N1-methyladenine and N3-methylcytosine from DNA
- PMID: 17396151
- PMCID: PMC1852788
- DOI: 10.1038/sj.emboj.7601662
Structural basis for enzymatic excision of N1-methyladenine and N3-methylcytosine from DNA
Abstract
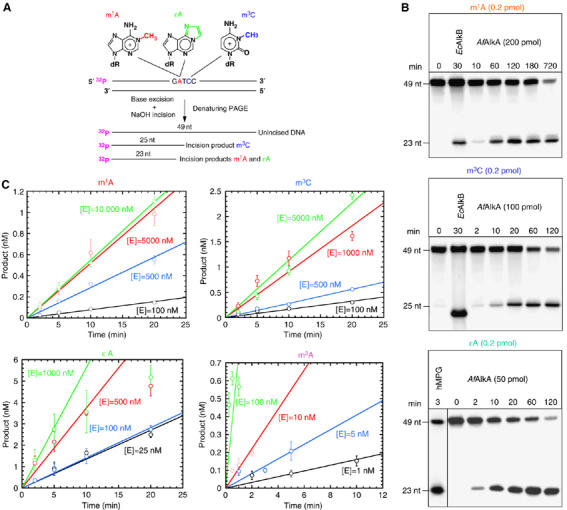
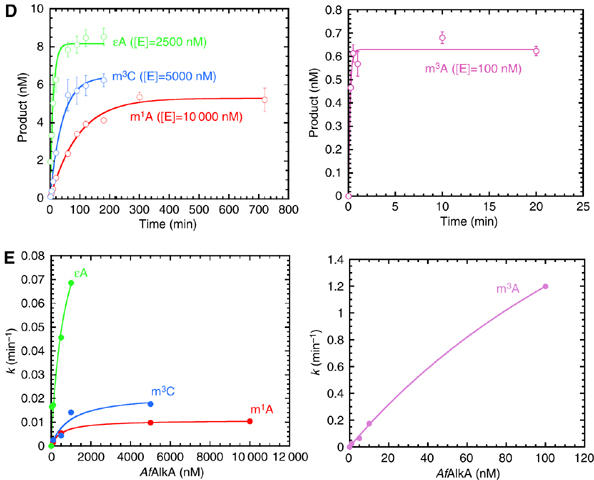
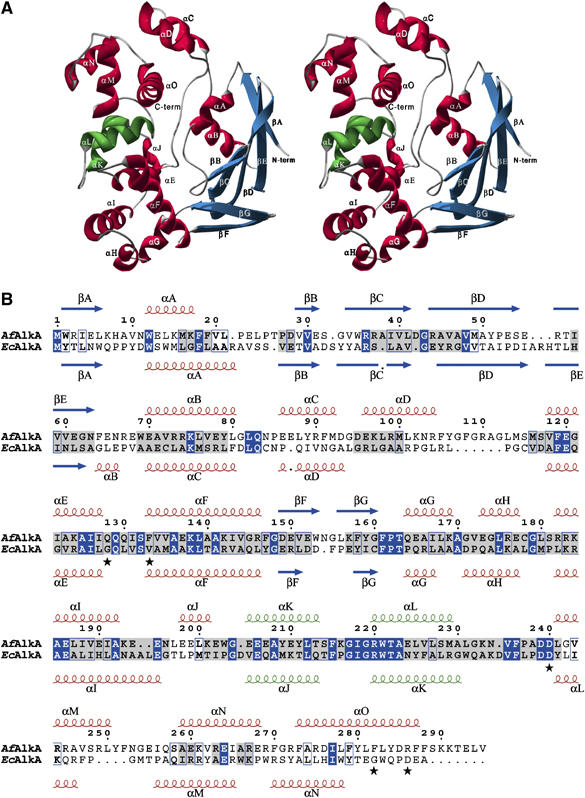
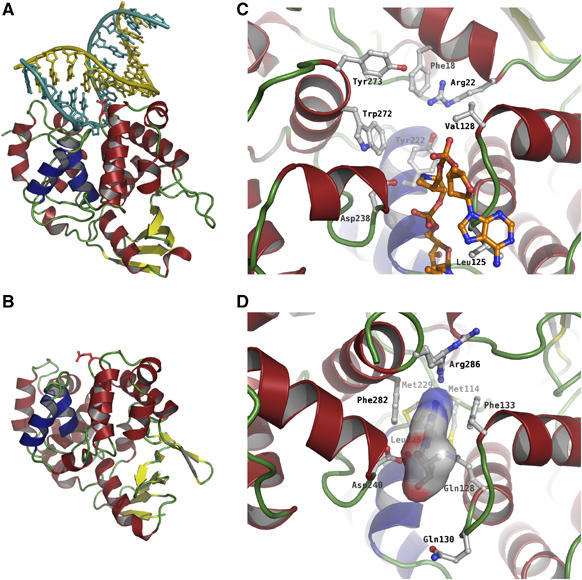
N(1)-methyladenine (m(1)A) and N(3)-methylcytosine (m(3)C) are major toxic and mutagenic lesions induced by alkylation in single-stranded DNA. In bacteria and mammals, m(1)A and m(3)C were recently shown to be repaired by AlkB-mediated oxidative demethylation, a direct DNA damage reversal mechanism. No AlkB gene homologues have been identified in Archaea. We report that m(1)A and m(3)C are repaired by the AfAlkA base excision repair glycosylase of Archaeoglobus fulgidus, suggesting a different repair mechanism for these lesions in the third domain of life. In addition, AfAlkA was found to effect a robust excision of 1,N(6)-ethenoadenine. We present a high-resolution crystal structure of AfAlkA, which, together with the characterization of several site-directed mutants, forms a molecular rationalization for the newly discovered base excision activity.
Figures


References
-
- Aas PA, Otterlei M, Falnes PØ, Vågbø CB, Skorpen F, Akbari M, Sundheim O, Bjørås M, Slupphaug G, Seeberg E, Krokan HE (2003) Human and bacterial oxidative demethylases repair alkylation damage in both RNA and DNA. Nature 421: 859–863 - PubMed
-
- Abner CW, Lau AY, Ellenberger T, Bloom LB (2001) Base excision and DNA binding activities of human alkyladenine DNA glycosylase are sensitive to the base paired with a lesion. J Biol Chem 276: 13379–13387 - PubMed
-
- Barrows LR, Magee PN (1982) Nonenzymatic methylation of DNA by S-adenosylmethionine in vitro. Carcinogenesis 3: 349–351 - PubMed
-
- Beranek DT (1990) Distribution of methyl and ethyl adducts following alkylation with monofunctional alkylating agents. Mutat Res 231: 11–30 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
