

Regulation of gluconeogenesis by Krüppel-like factor 15
- PMID: 17403374
- PMCID: PMC1892530
- DOI: 10.1016/j.cmet.2007.03.002
Regulation of gluconeogenesis by Krüppel-like factor 15
Abstract
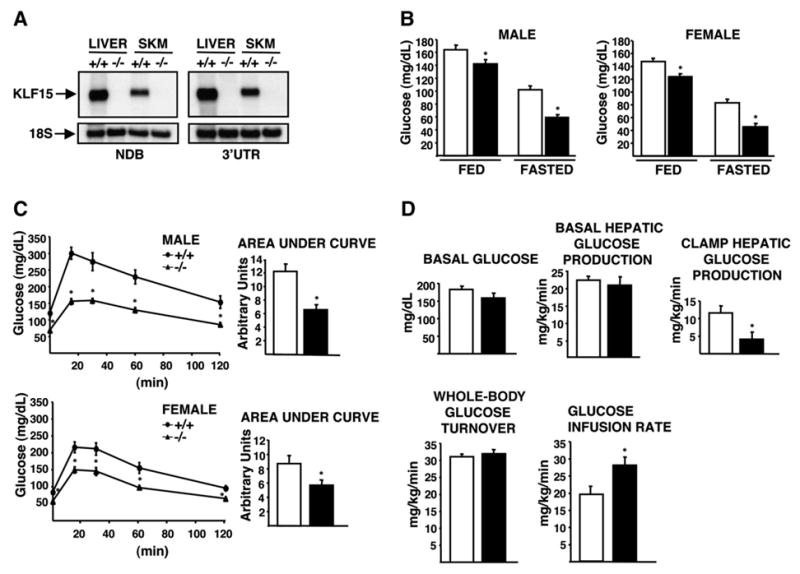
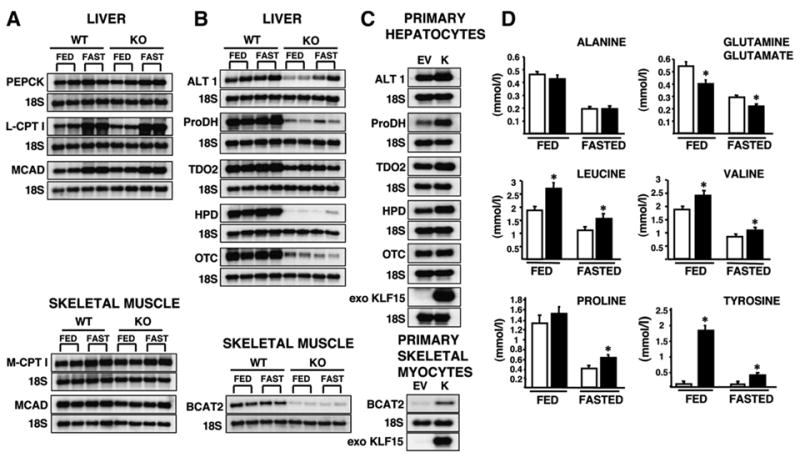
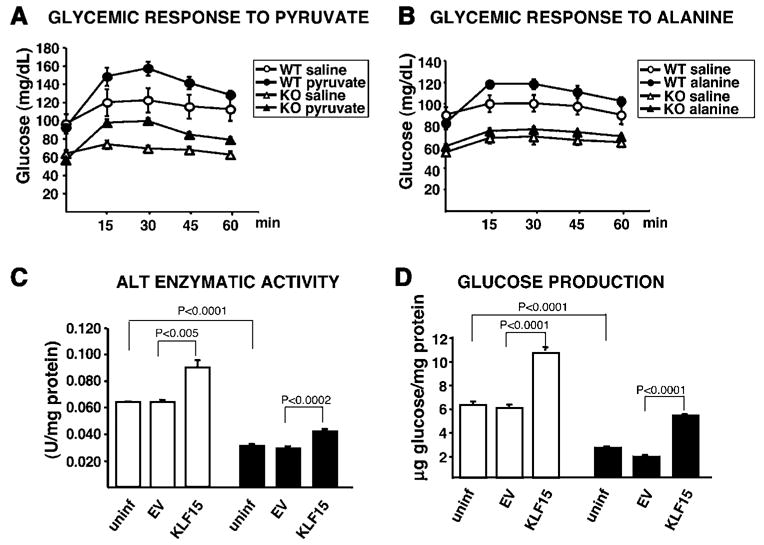
In the postabsorptive state, certain tissues, including the brain, require glucose as the sole source of energy. After an overnight fast, hepatic glycogen stores are depleted, and gluconeogenesis becomes essential for preventing life-threatening hypoglycemia. Mice with a targeted deletion of KLF15, a member of the Krüppel-like family of transcription factors, display severe hypoglycemia after an overnight (18 hr) fast. We provide evidence that defective amino acid catabolism promotes the development of fasting hypoglycemia in KLF15-/- mice by limiting gluconeogenic substrate availability. KLF15-/- liver and skeletal muscle show markedly reduced mRNA expression of amino acid-degrading enzymes. Furthermore, the enzymatic activity of alanine aminotransferase (ALT), which converts the critical gluconeogenic amino acid alanine into pyruvate, is decreased (approximately 50%) in KLF15-/- hepatocytes. Consistent with this observation, intraperitoneal injection of pyruvate, but not alanine, rescues fasting hypoglycemia in KLF15-/- mice. We conclude that KLF15 plays an important role in the regulation of gluconeogenesis.
Figures
References
-
- Bieker JJ. Kruppel-like factors: three fingers in many pies. J Biol Chem. 2001;276:34355–34358. - PubMed
-
- Burelle Y, Fillipi C, Peronnet F, Leverve X. Mechanisms of increased gluconeogenesis from alanine in rat isolated hepatocytes after endurance training. Am J Physiol Endocrinol Metab. 2000;278:E35–E42. - PubMed
-
- Chochinov R, Perlman K, Moorhouse J. Circulating alanine production and disposal in healthy subjects. Diabetes. 1978;27:287–295. - PubMed
-
- Connett RJ, Honig CR, Gayeski TE, Brooks GA. Defining hypoxia: a systems view of VO2, glycolysis, energetics, and intracellular PO2. J Appl Physiol. 1990;68:833–842. - PubMed
-
- Consoli A, Nurjhan N, Reilly JJ, Jr, Bier DM, Gerich JE. Contribution of liver and skeletal muscle to alanine and lactate metabolism in humans. Am J Physiol Endocrinol Metab. 1990;259:E677–E684. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R01 HL072952/HL/NHLBI NIH HHS/United States
- F32 HL077052/HL/NHLBI NIH HHS/United States
- P01 HL048743/HL/NHLBI NIH HHS/United States
- R01 DK36836/DK/NIDDK NIH HHS/United States
- U24 DK59635/DK/NIDDK NIH HHS/United States
- HL76754/HL/NHLBI NIH HHS/United States
- R01 HL076754/HL/NHLBI NIH HHS/United States
- R01 HL075427/HL/NHLBI NIH HHS/United States
- U24 DK059635/DK/NIDDK NIH HHS/United States
- P30 DK036836/DK/NIDDK NIH HHS/United States
- K01 DK064950/DK/NIDDK NIH HHS/United States
- HL48743/HL/NHLBI NIH HHS/United States
- HL72952/HL/NHLBI NIH HHS/United States
- P30 DK057521/DK/NIDDK NIH HHS/United States
- R01 DK043051/DK/NIDDK NIH HHS/United States
- DK57521/DK/NIDDK NIH HHS/United States
- DK064950/DK/NIDDK NIH HHS/United States
- R01 DK080756/DK/NIDDK NIH HHS/United States
- HL75427/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
