

Two small regulatory RNAs establish opposing fates of a developmental axis
- PMID: 17403777
- PMCID: PMC1838527
- DOI: 10.1101/gad.1528607
Two small regulatory RNAs establish opposing fates of a developmental axis
Abstract
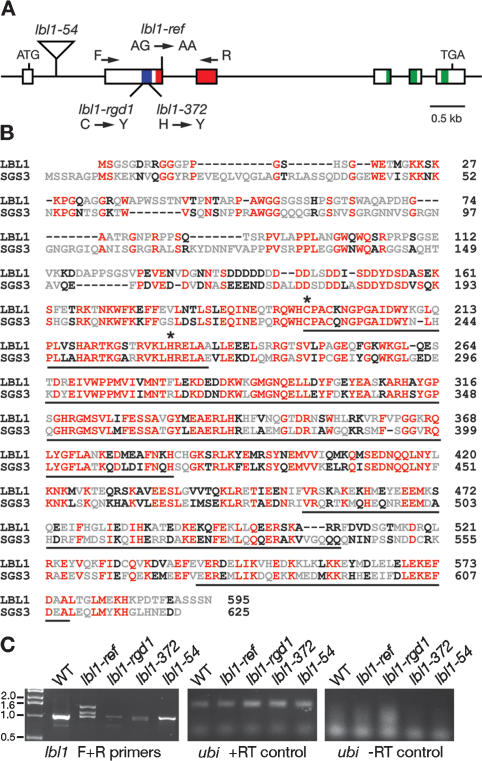
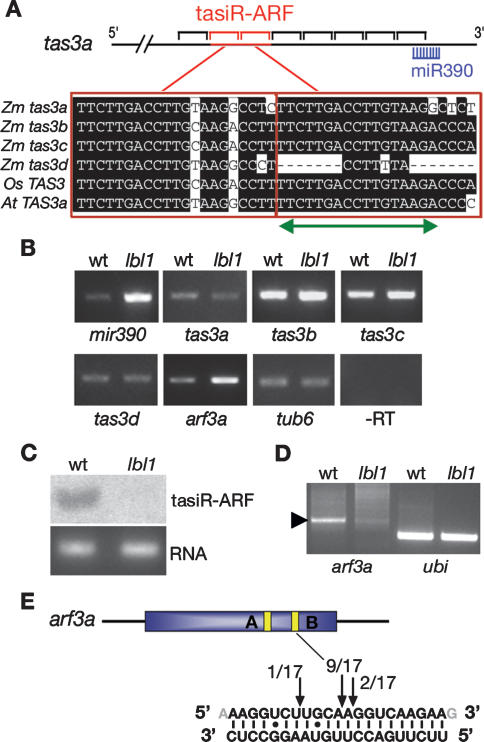
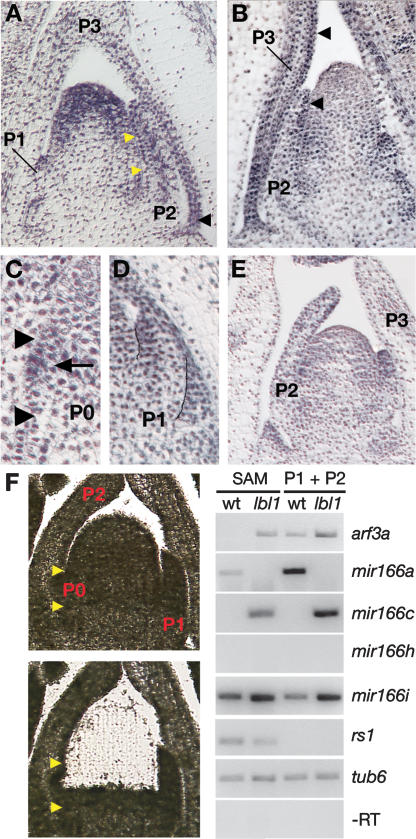
Small RNAs are important regulators of gene expression. In maize, adaxial/abaxial (dorsoventral) leaf polarity is established by an abaxial gradient of microRNA166 (miR166), which spatially restricts the expression domain of class III homeodomain leucine zipper (HD-ZIPIII) transcription factors that specify adaxial/upper fate. Here, we show that leafbladeless1 encodes a key component in the trans-acting small interfering RNA (ta-siRNA) biogenesis pathway that acts on the adaxial side of developing leaves and demarcates the domains of hd-zipIII and miR166 accumulation. Our findings indicate that tasiR-ARF, a ta-siRNA, and miR166 establish opposing domains along the adaxial-abaxial axis, thus revealing a novel mechanism of pattern formation.
Figures

References
-
- Adenot X., Elmayan T., Lauressergues D., Boutet S., Bouche N., Gasciolli V., Vaucheret H., Elmayan T., Lauressergues D., Boutet S., Bouche N., Gasciolli V., Vaucheret H., Lauressergues D., Boutet S., Bouche N., Gasciolli V., Vaucheret H., Boutet S., Bouche N., Gasciolli V., Vaucheret H., Bouche N., Gasciolli V., Vaucheret H., Gasciolli V., Vaucheret H., Vaucheret H. DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Curr. Biol. 2006;16:927–932. - PubMed
-
- Allen E., Xie Z., Gustafson A., Carrington J., Xie Z., Gustafson A., Carrington J., Gustafson A., Carrington J., Carrington J. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell. 2005;121:207–221. - PubMed
-
- Alvarez-Garcia I., Miska E., Miska E. MicroRNA functions in animal development and human disease. Development. 2005;132:4653–4662. - PubMed
-
- Bartel D. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
-
- Dunoyer P., Himber C., Voinnet O., Himber C., Voinnet O., Voinnet O. DICER-LIKE 4 is required for RNA interference and produces the 21-nucleotide small interfering RNA component of the plant cell-to-cell silencing signal. Nat. Genet. 2005;37:1356–1360. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources