

Eos, MITF, and PU.1 recruit corepressors to osteoclast-specific genes in committed myeloid progenitors
- PMID: 17403896
- PMCID: PMC1900027
- DOI: 10.1128/MCB.01839-06
Eos, MITF, and PU.1 recruit corepressors to osteoclast-specific genes in committed myeloid progenitors
Abstract
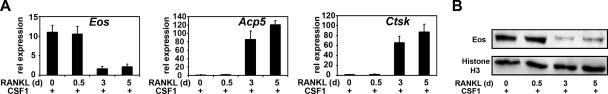
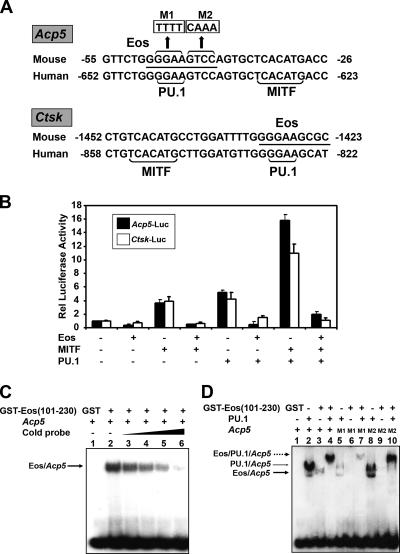
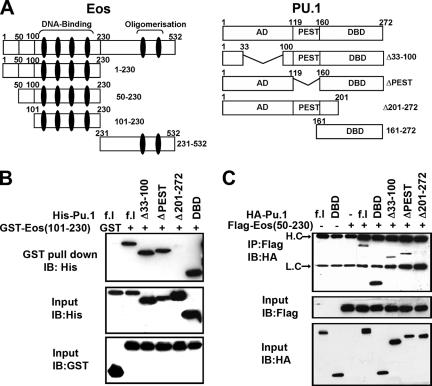
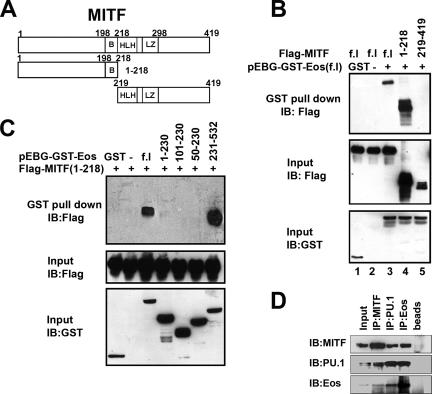
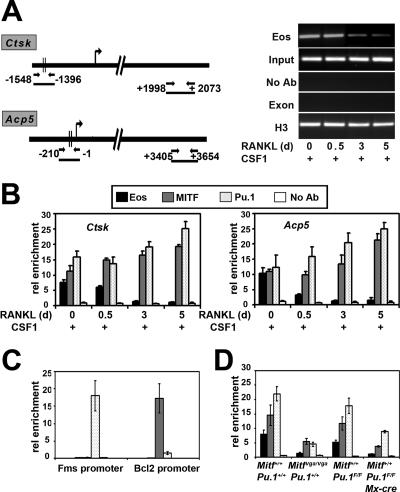
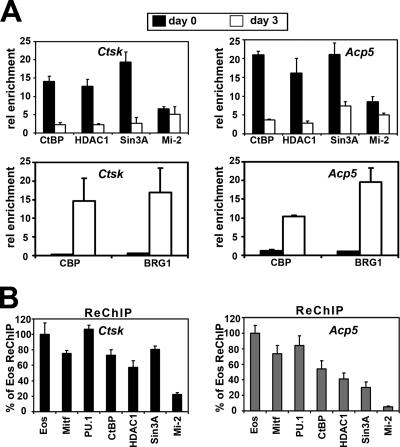
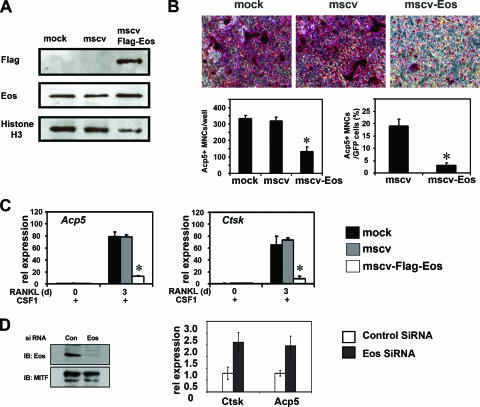
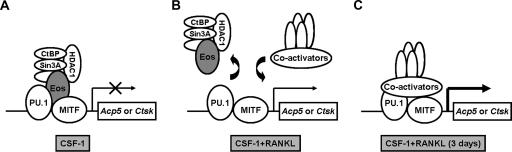
Transcription factors MITF and PU.1 collaborate to increase expression of target genes like cathepsin K (Ctsk) and acid phosphatase 5 (Acp5) during osteoclast differentiation. We show that these factors can also repress transcription of target genes in committed myeloid precursors capable of forming either macrophages or osteoclasts. The direct interaction of MITF and PU.1 with the zinc finger protein Eos, an Ikaros family member, was necessary for repression of Ctsk and Acp5. Eos formed a complex with MITF and PU.1 at target gene promoters and suppressed transcription through recruitment of corepressors CtBP (C-terminal binding protein) and Sin3A, but during osteoclast differentiation, Eos association with Ctsk and Acp5 promoters was significantly decreased. Subsequently, MITF and PU.1 recruited coactivators to these target genes, resulting in robust expression of target genes. Overexpression of Eos in bone marrow-derived precursors disrupted osteoclast differentiation and selectively repressed transcription of MITF/PU.1 targets, while small interfering RNA knockdown of Eos resulted in increased basal expression of Ctsk and Acp5. This work provides a mechanism to account for the modulation of MITF and PU.1 activity in committed myeloid progenitors prior to the initiation of osteoclast differentiation in response to the appropriate extracellular signals.
Figures
Similar articles
-
MITF and PU.1 recruit p38 MAPK and NFATc1 to target genes during osteoclast differentiation.J Biol Chem. 2007 May 25;282(21):15921-9. doi: 10.1074/jbc.M609723200. Epub 2007 Apr 2. J Biol Chem. 2007. PMID: 17403683
-
Defective co-activator recruitment in osteoclasts from microphthalmia-oak ridge mutant mice.J Cell Physiol. 2009 Jul;220(1):230-7. doi: 10.1002/jcp.21755. J Cell Physiol. 2009. PMID: 19288495 Free PMC article.
-
The multifunctional protein fused in sarcoma (FUS) is a coactivator of microphthalmia-associated transcription factor (MITF).J Biol Chem. 2014 Jan 3;289(1):326-34. doi: 10.1074/jbc.M113.493874. Epub 2013 Nov 20. J Biol Chem. 2014. PMID: 24257758 Free PMC article.
-
Mechanisms of Action of Hematopoietic Transcription Factor PU.1 in Initiation of T-Cell Development.Front Immunol. 2019 Feb 20;10:228. doi: 10.3389/fimmu.2019.00228. eCollection 2019. Front Immunol. 2019. PMID: 30842770 Free PMC article. Review.
-
The Yin and Yang of Sox proteins: Activation and repression in development and disease.J Neurosci Res. 2009 Nov 15;87(15):3277-87. doi: 10.1002/jnr.22128. J Neurosci Res. 2009. PMID: 19437544 Free PMC article. Review.
Cited by
-
EZH2 Supports Osteoclast Differentiation and Bone Resorption Via Epigenetic and Cytoplasmic Targets.J Bone Miner Res. 2020 Jan;35(1):181-195. doi: 10.1002/jbmr.3863. Epub 2019 Oct 23. J Bone Miner Res. 2020. PMID: 31487061 Free PMC article.
-
CSF1-ETS2-induced microRNA in myeloid cells promote metastatic tumor growth.Oncogene. 2015 Jul;34(28):3651-61. doi: 10.1038/onc.2014.294. Epub 2014 Sep 22. Oncogene. 2015. PMID: 25241894 Free PMC article.
-
Negative regulation of osteoclastogenesis and bone resorption by cytokines and transcriptional repressors.Arthritis Res Ther. 2011 Jul 28;13(4):234. doi: 10.1186/ar3379. Arthritis Res Ther. 2011. PMID: 21861861 Free PMC article. Review.
-
Targeted analysis of four breeds narrows equine Multiple Congenital Ocular Anomalies locus to 208 kilobases.Mamm Genome. 2011 Jun;22(5-6):353-60. doi: 10.1007/s00335-011-9325-7. Epub 2011 Apr 5. Mamm Genome. 2011. PMID: 21465164 Free PMC article.
-
Bone Remodeling: Histone Modifications as Fate Determinants of Bone Cell Differentiation.Int J Mol Sci. 2019 Jun 27;20(13):3147. doi: 10.3390/ijms20133147. Int J Mol Sci. 2019. PMID: 31252653 Free PMC article. Review.
References
-
- Boyle, W. J., W. S. Simonet, and D. L. Lacey. 2003. Osteoclast differentiation and activation. Nature 423:337-342. - PubMed
-
- de la Serna, I. L., Y. Ohkawa, C. Higashi, C. Dutta, J. Osias, N. Kommajosyula, T. Tachibana, and A. N. Imbalzano. 2006. The microphthalmia-associated transcription factor requires SWI/SNF enzymes to activate melanocyte-specific genes. J. Biol. Chem. 281:20233-20241. - PubMed
-
- Georgopoulos, K., M. Bigby, J. H. Wang, A. Molnar, P. Wu, S. Winandy, and A. Sharpe. 1994. The Ikaros gene is required for the development of all lymphoid lineages. Cell 79:143-156. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases