

Conformational modification of serpins transforms leukocyte elastase inhibitor into an endonuclease involved in apoptosis
- PMID: 17403905
- PMCID: PMC1900025
- DOI: 10.1128/MCB.01959-06
Conformational modification of serpins transforms leukocyte elastase inhibitor into an endonuclease involved in apoptosis
Abstract
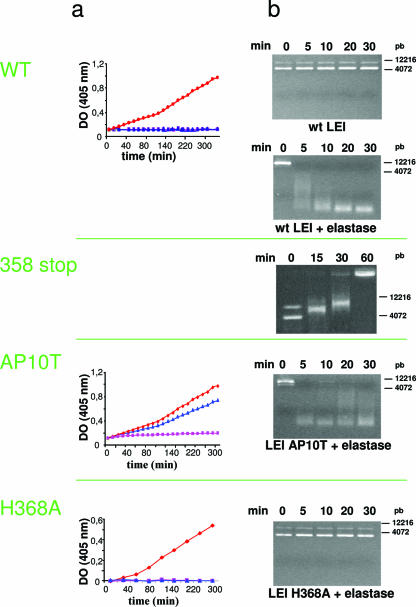
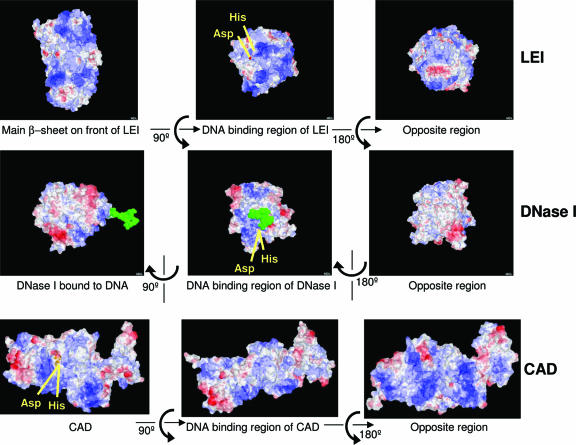
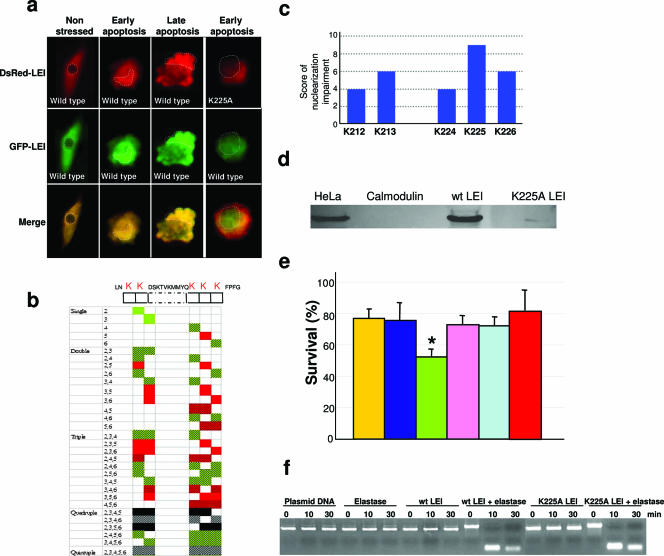
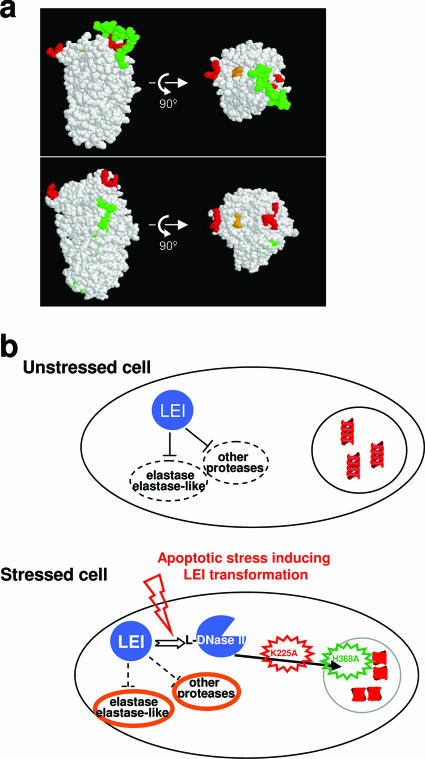
The best-characterized biochemical feature of apoptosis is degradation of genomic DNA into oligonucleosomes. The endonuclease responsible for DNA degradation in caspase-dependent apoptosis is caspase-activated DNase. In caspase-independent apoptosis, different endonucleases may be activated according to the cell line and the original insult. Among the known effectors of caspase-independent cell death, L-DNase II (LEI [leukocyte elastase inhibitor]-derived DNase II) has been previously characterized by our laboratory. We have thus shown that this endonuclease derives from the serpin superfamily member LEI by posttranslational modification (A. Torriglia, P. Perani, J. Y. Brossas, E. Chaudun, J. Treton, Y. Courtois, and M. F. Counis, Mol. Cell. Biol. 18:3612-3619, 1998). In this work, we assessed the molecular mechanism involved in the change in the enzymatic activity of this molecule from an antiprotease to an endonuclease. We report that the cleavage of LEI by elastase at its reactive center loop abolishes its antiprotease activity and leads to a conformational modification that exposes an endonuclease active site and a nuclear localization signal. This represents a novel molecular mechanism for a complete functional conversion induced by changing the conformation of a serpin. We also show that this molecular transformation affects cellular fate and that both endonuclease activity and nuclear translocation of L-DNase II are needed to induce cell death.
Figures

Similar articles
-
Nuclear export of LEI/L-DNase II by Crm1 is essential for cell survival.Biochim Biophys Acta. 2008 Jun;1783(6):1068-75. doi: 10.1016/j.bbamcr.2008.02.012. Epub 2008 Feb 23. Biochim Biophys Acta. 2008. PMID: 18342633
-
Leukocyte elastase inhibitor: a new regulator of PARP-1.Ann N Y Acad Sci. 2009 Aug;1171:25-31. doi: 10.1111/j.1749-6632.2009.04701.x. Ann N Y Acad Sci. 2009. PMID: 19723034
-
Apoptosis induced by Na+/H+ antiport inhibition activates the LEI/L-DNase II pathway.Cell Death Differ. 2003 May;10(5):548-57. doi: 10.1038/sj.cdd.4401195. Cell Death Differ. 2003. PMID: 12728253
-
Acid DNases and their interest among apoptotic endonucleases.Biochimie. 2006 Dec;88(12):1851-8. doi: 10.1016/j.biochi.2006.07.008. Epub 2006 Aug 8. Biochimie. 2006. PMID: 16989934 Review.
-
Molecular mechanism of L-DNase II activation and function as a molecular switch in apoptosis.Biochem Pharmacol. 2008 Dec 1;76(11):1490-502. doi: 10.1016/j.bcp.2008.07.039. Epub 2008 Aug 9. Biochem Pharmacol. 2008. PMID: 18761000 Review.
Cited by
-
Inhibitors of Serine Proteases in Regulating the Production and Function of Neutrophil Extracellular Traps.Front Immunol. 2016 Jun 30;7:261. doi: 10.3389/fimmu.2016.00261. eCollection 2016. Front Immunol. 2016. PMID: 27446090 Free PMC article. Review.
-
Schistosoma japonicum serine protease inhibitor increases endothelial barrier function.Int J Clin Exp Pathol. 2017 Jul 1;10(7):7312-7324. eCollection 2017. Int J Clin Exp Pathol. 2017. PMID: 31966571 Free PMC article.
-
The hidden side of SERPINB1/Leukocyte Elastase Inhibitor.Semin Cell Dev Biol. 2017 Feb;62:178-186. doi: 10.1016/j.semcdb.2016.07.010. Epub 2016 Jul 12. Semin Cell Dev Biol. 2017. PMID: 27422329 Free PMC article. Review.
-
Effects of white light-emitting diode (LED) exposure on retinal pigment epithelium in vivo.J Cell Mol Med. 2017 Dec;21(12):3453-3466. doi: 10.1111/jcmm.13255. Epub 2017 Jun 29. J Cell Mol Med. 2017. PMID: 28661040 Free PMC article.
-
The activation of the atypical PKC zeta in light-induced retinal degeneration and its involvement in L-DNase II control.J Cell Mol Med. 2015 Jul;19(7):1646-55. doi: 10.1111/jcmm.12539. Epub 2015 Mar 17. J Cell Mol Med. 2015. PMID: 25781645 Free PMC article.
References
-
- Altairac, S., S. C. Wright, Y. Courtois, and A. Torriglia. 2003. L-DNase II activation by the 24 kDa apoptotic protease (AP24) in TNFα-induced apoptosis. Cell Death Differ. 10:1109-1111. - PubMed
-
- Altairac, S., S. Zeggai, P. Perani, Y. Courtois, and A. Torriglia. 2003. Apoptosis induced by Na+/H+ antiport inhibition activates the LEI/L-DNase II pathway. Cell Death Differ. 10:548-557. - PubMed
-
- Barry, M. A., and A. Eastman. 1993. Identification of deoxyribonuclease II as an endonuclease involved in apoptosis. Arch. Biochem. Biophys. 300:440-450. - PubMed
-
- Belmokhtar, C. A., J. Hillion, C. Dudognon, S. Fiorentino, M. Flexor, M. Lanotte, and E. Segal-Bendirdjian. 2003. Apoptosome-independent pathway for apoptosis. Biochemical analysis of APAF-1 defects and biological outcomes. J. Biol. Chem. 278:29571-29580. - PubMed
-
- Belmokhtar, C. A., A. Torriglia, M. F. Counis, Y. Courtois, A. Jacquemin-Sablon, and E. Segal-Bendirdjian. 2000. Nuclear translocation of a leukocyte elastase inhibitor/elastase complex during staurosporine-induced apoptosis: role in the generation of nuclear L-DNase II activity. Exp. Cell Res. 254:99-109. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources