

A role for a novel centrosome cycle in asymmetric cell division
- PMID: 17403931
- PMCID: PMC2064101
- DOI: 10.1083/jcb.200612140
A role for a novel centrosome cycle in asymmetric cell division
Abstract
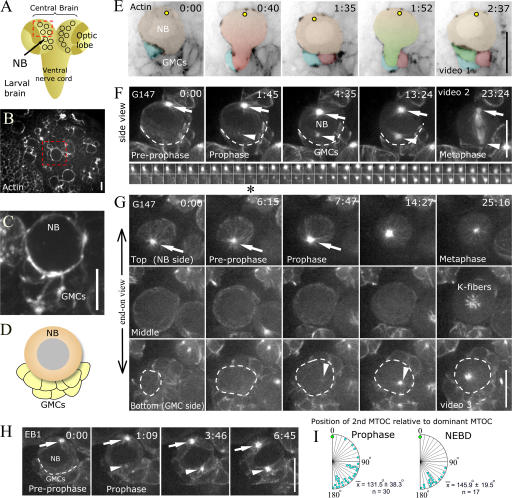
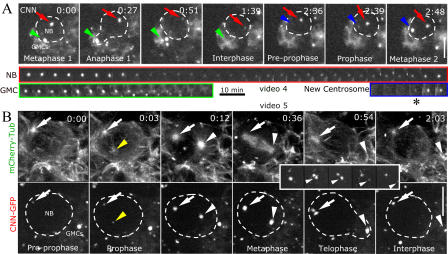
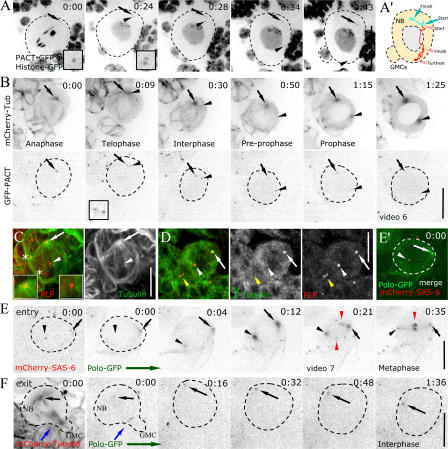
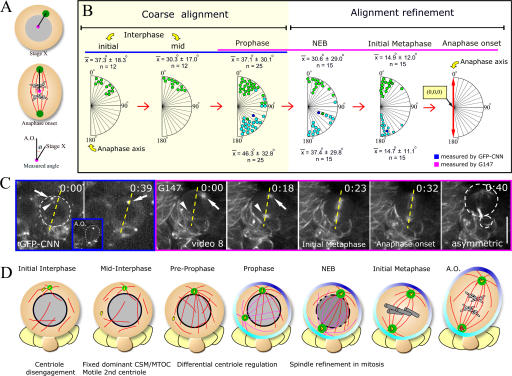
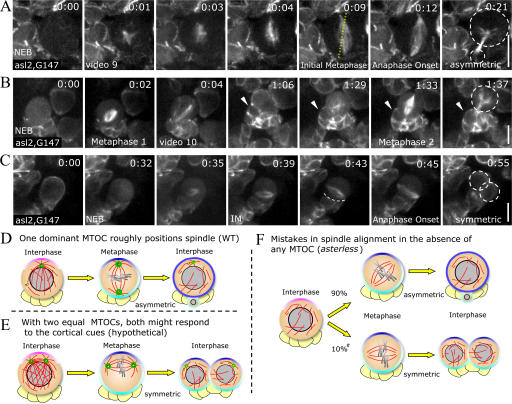
Tissue stem cells play a key role in tissue maintenance. Drosophila melanogaster central brain neuroblasts are excellent models for stem cell asymmetric division. Earlier work showed that their mitotic spindle orientation is established before spindle formation. We investigated the mechanism by which this occurs, revealing a novel centrosome cycle. In interphase, the two centrioles separate, but only one is active, retaining pericentriolar material and forming a "dominant centrosome." This centrosome acts as a microtubule organizing center (MTOC) and remains stationary, forming one pole of the future spindle. The second centriole is inactive and moves to the opposite side of the cell before being activated as a centrosome/MTOC. This is accompanied by asymmetric localization of Polo kinase, a key centrosome regulator. Disruption of centrosomes disrupts the high fidelity of asymmetric division. We propose a two-step mechanism to ensure faithful spindle positioning: the novel centrosome cycle produces a single interphase MTOC, coarsely aligning the spindle, and spindle-cortex interactions refine this alignment.
Figures
References
-
- Akong, K., B.M. McCartney, and M. Peifer. 2002. Drosophila APC2 and APC1 have overlapping roles in the larval brain despite their distinct intracellular localizations. Dev. Biol. 250:71–90. - PubMed
-
- Alberts, B., A. Johnson, J. Lewis, M. Raff, K. Roberts, and P. Walter. 2002. Molecular biology of the cell. Garland Science, New York. 1616 pp.
-
- Basto, R., J. Lau, T. Vinogradova, A. Gardiol, C.G. Woods, A. Khodjakov, and J.W. Raff. 2006. Flies without centrioles. Cell. 125:1375–1386. - PubMed
-
- Callaini, G., and M.G. Riparbelli. 1990. Centriole and centrosome cycle in the early Drosophila embryo. J. Cell Sci. 97:539–543. - PubMed
-
- Ceron, J., C. Gonzalez, and F.J. Tejedor. 2001. Patterns of cell division and expression of asymmetric cell fate determinants in postembryonic neuroblast lineages of Drosophila. Dev. Biol. 230:125–138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
