

c-Met is essential for wound healing in the skin
- PMID: 17403932
- PMCID: PMC2064119
- DOI: 10.1083/jcb.200701086
c-Met is essential for wound healing in the skin
Abstract
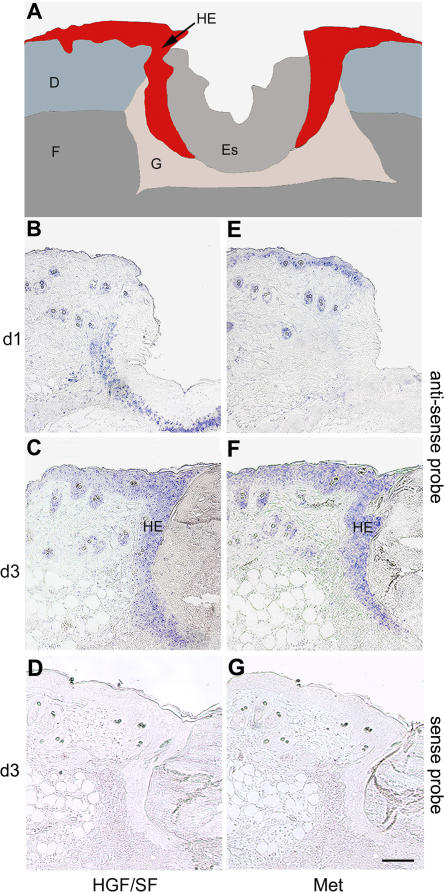
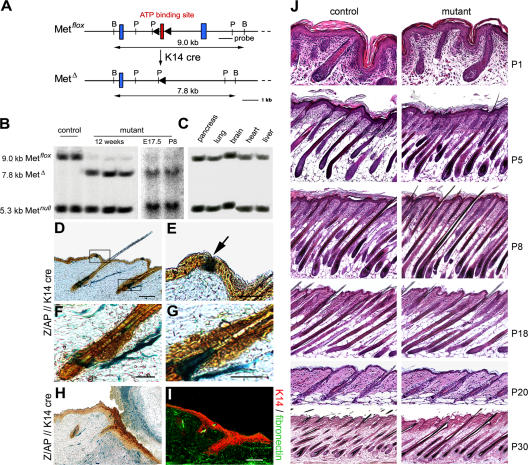
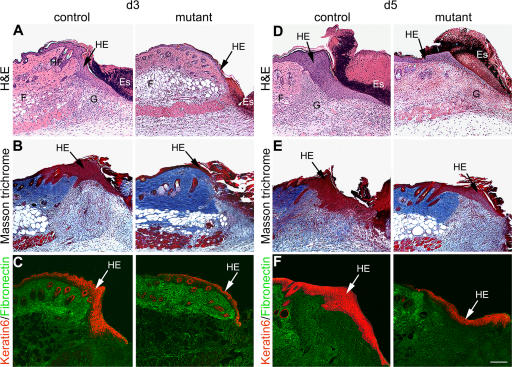
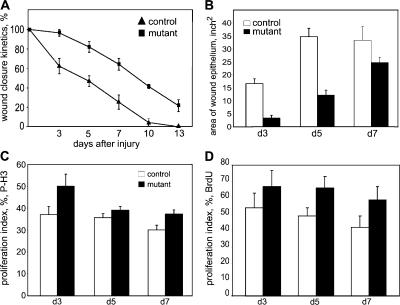
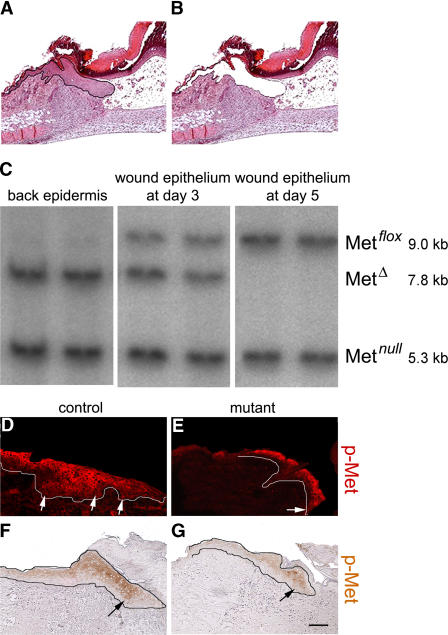
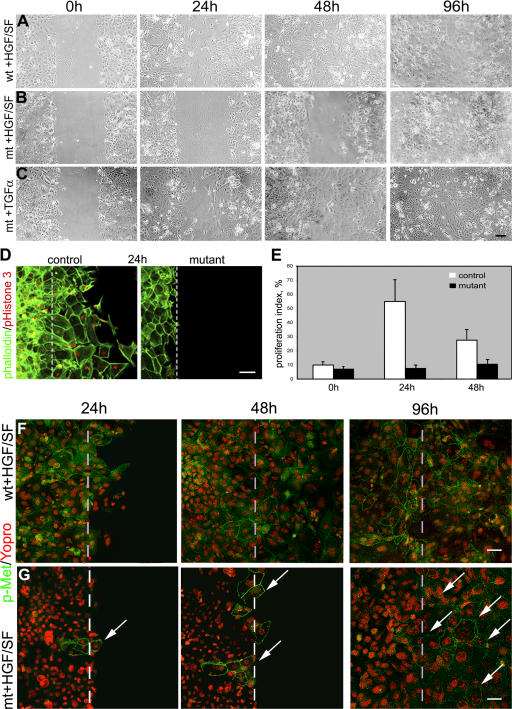
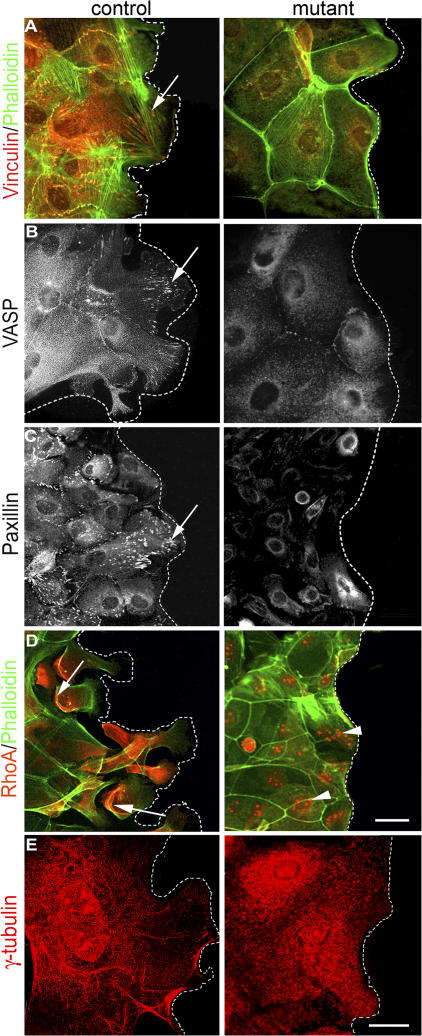
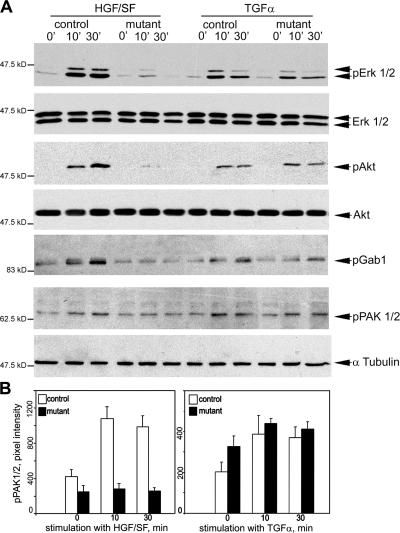
Wound healing of the skin is a crucial regenerative process in adult mammals. We examined wound healing in conditional mutant mice, in which the c-Met gene that encodes the receptor of hepatocyte growth factor/scatter factor was mutated in the epidermis by cre recombinase. c-Met-deficient keratinocytes were unable to contribute to the reepithelialization of skin wounds. In conditional c-Met mutant mice, wound closure was slightly attenuated, but occurred exclusively by a few (5%) keratinocytes that had escaped recombination. This demonstrates that the wound process selected and amplified residual cells that express a functional c-Met receptor. We also cultured primary keratinocytes from the skin of conditional c-Met mutant mice and examined them in scratch wound assays. Again, closure of scratch wounds occurred by the few remaining c-Met-positive cells. Our data show that c-Met signaling not only controls cell growth and migration during embryogenesis but is also essential for the generation of the hyperproliferative epithelium in skin wounds, and thus for a fundamental regenerative process in the adult.
Figures
References
-
- Amendt, C., A. Mann, P. Schirmacher, and M. Blessing. 2002. Resistance of keratinocytes to TGFbeta-mediated growth restriction and apoptosis induction accelerates re-epithelialization in skin wounds. J. Cell Sci. 115:2189–2198. - PubMed
-
- Ashcroft, G.S., X. Yang, A.B. Glick, M. Weinstein, J.L. Letterio, D.E. Mizel, M. Anzano, T. Greenwell-Wild, S.M. Wahl, C. Deng, and A.B. Roberts. 1999. Mice lacking Smad3 show accelerated wound healing and an impaired local inflammatory response. Nat. Cell Biol. 1:260–266. - PubMed
-
- Bevan, D., E. Gherardi, T.P. Fan, D. Edwards, and R. Warn. 2004. Diverse and potent activities of HGF/SF in skin wound repair. J. Pathol. 203:831–838. - PubMed
-
- Birchmeier, C., and E. Gherardi. 1998. Developmental roles of HGF/SF and its receptor, the c-Met tyrosine kinase. Trends Cell Biol. 8:404–410. - PubMed
-
- Birchmeier, C., W. Birchmeier, E. Gherardi, and G.F. Vande Woude. 2003. Met, metastasis, motility and more. Nat. Rev. Mol. Cell Biol. 4:915–925. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
