

Onset rivalry: brief presentation isolates an early independent phase of perceptual competition
- PMID: 17406667
- PMCID: PMC1828625
- DOI: 10.1371/journal.pone.0000343
Onset rivalry: brief presentation isolates an early independent phase of perceptual competition
Abstract
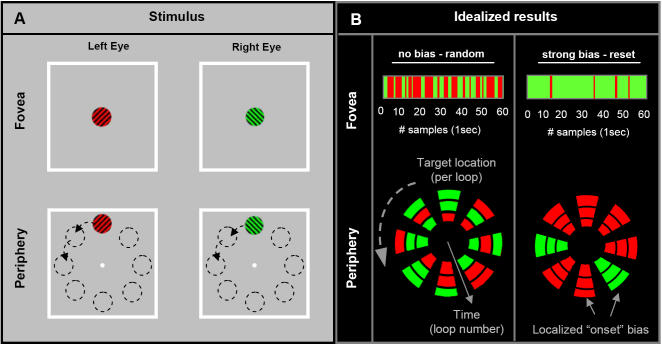
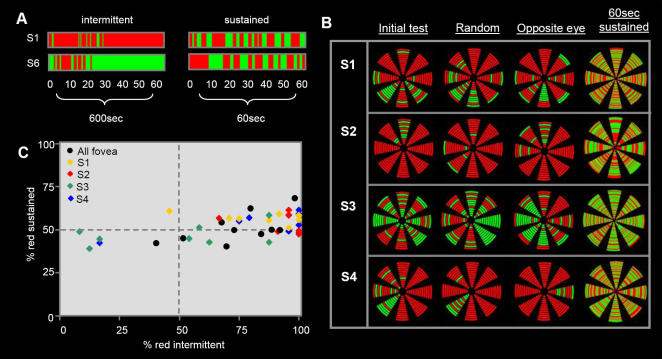
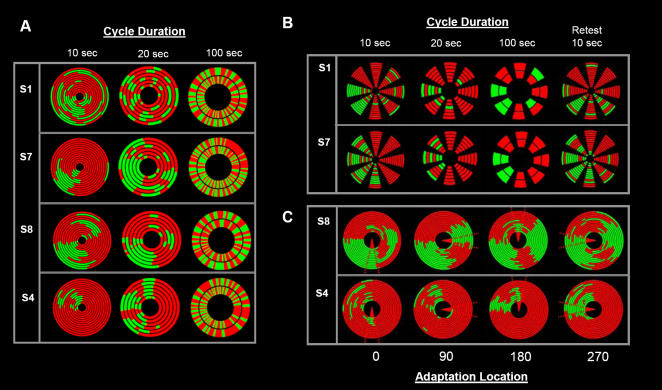
When the left and right eyes are simultaneously presented with different images, observers typically report exclusive awareness of only one image. This phenomenon is termed binocular rivalry, reflecting the fact that the dominant image alternates every few seconds in a cycle of perceptual competition that continues indefinitely. Despite the apparent continuity in perceptual switching, we now demonstrate that the initial "onset" period is fundamentally different to all subsequent rivalry epochs. Using brief intermittent presentations, rivalry dominance shows strong biases such that the same target is perceived with each successive stimulus onset. These biases remain consistent within any given location, but vary across the visual field in a distribution that is stable over multiple weeks but highly idiosyncratic across observers. If the presentation exceeds approximately 1sec at any location, however, the very different and much more balanced alternations of sustained binocular rivalry become apparent. These powerful onset biases are observed with brief intermittent presentations at a single location or with continual smooth motion of the targets. Periods of adaptation to one of the rivaling targets induced local switches in dominance to the non-adapted target. However, these effects were generally limited to the spatial site of adaptation and had less influence over each subsequent cycle of the target. We conclude that onset rivalry is independent of sustained rivalry and cannot be explained by local regions of monocular dominance or memory of past perceptual history, but rather reflects low-level, spatially localized factors that are stable over periods of weeks. These findings suggest that brief presentation paradigms are inappropriate for their current use in studies of the mechanisms underlying sustained rivalry. However, brief presentations are ideal for investigating early stages of perceptual competition.
Conflict of interest statement
Figures
References
-
- Wheatstone C. On some remarkable, and hitherto unobserved, phenomena of binocular vision. Philos Trans R Soc Lond. 1838;128:371–394. - PubMed
-
- Blake R, Logothetis N. Visual competition. Nat Rev Neurosci. 2002;3:13–21. - PubMed
-
- Maier A, Wilke M, Logothetis NK, Leopold AD. Perception of temporally interleaved ambiguous patterns. Curr Biol. 2003;13:1076–1085. - PubMed
-
- Carney T, Shadlen M, Switkes E. Parallel processing of motion and colour information. Nature. 1987;328:647–649. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
