

A role for plasmacytoid dendritic cells in the rapid IL-18-dependent activation of NK cells following HSV-1 infection
- PMID: 17407097
- PMCID: PMC2699043
- DOI: 10.1002/eji.200636362
A role for plasmacytoid dendritic cells in the rapid IL-18-dependent activation of NK cells following HSV-1 infection
Abstract
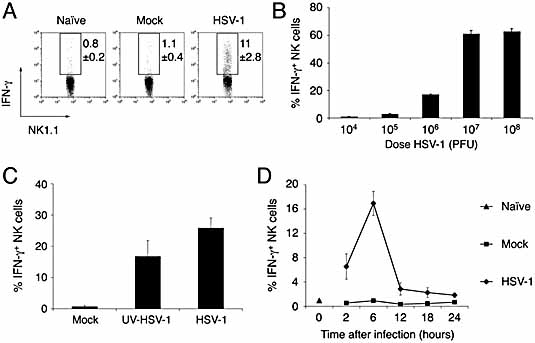
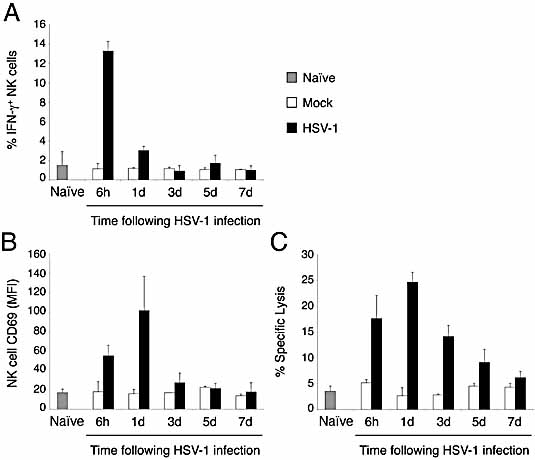
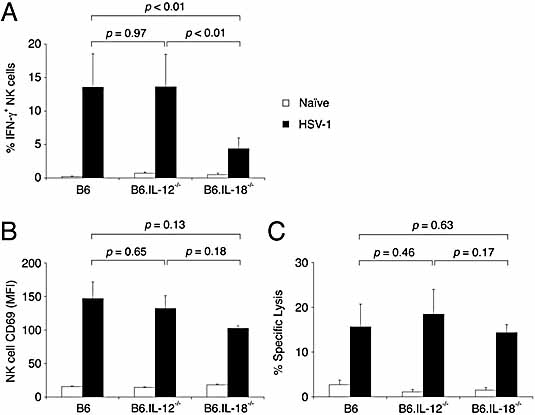
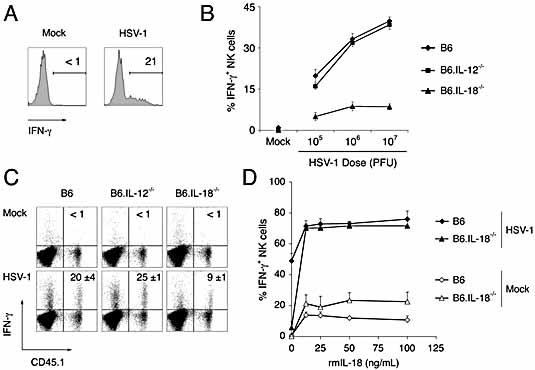
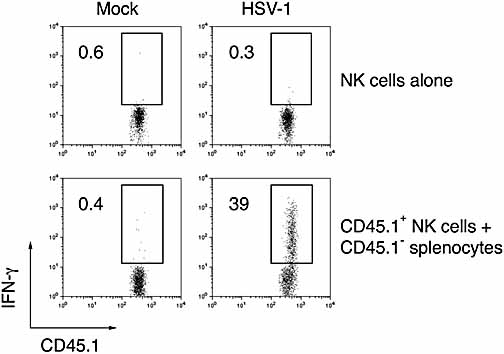
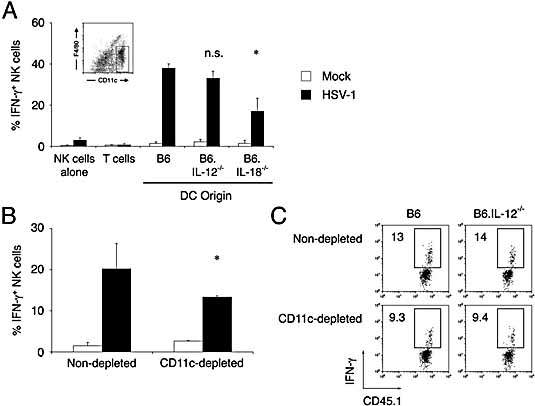
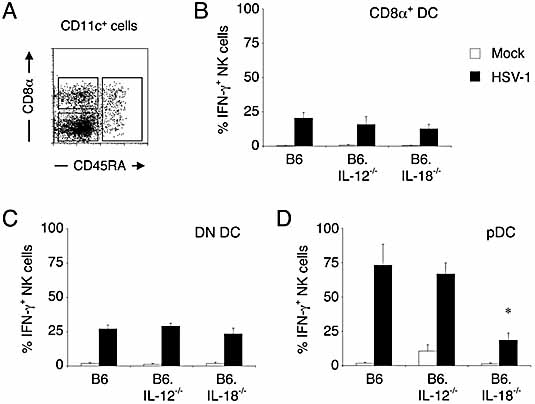
Natural killer (NK) cells play a crucial role in the initial response to viral infections but the mechanisms controlling their activation are unclear. We show a rapid and transient activation of NK cells that results in the production of IFN-gamma immediately following infection with herpes simplex virus type 1 (HSV-1). Activation of NK cells leading to synthesis of IFN-gamma was not mediated by a direct interaction with virus but required the presence of additional cell types and was largely dependent on the cytokine IL-18, but not IL-12. HSV-1-induced IFN-gamma expression by NK cells in vitro was impaired in spleen cultures depleted of CD11c(+) cells. Conversely, coculture of NK cells with virus-exposed conventional DC or plasmacytoid (p)DC restored the production of IFN-gamma, indicating that multiple DC subsets could mediate NK cell activation. While conventional DC populations stimulated NK cells independently of IL-18, they were less effective than pDC in promoting NK cell IFN-gamma expression. In contrast, the potent stimulation of NK cells by pDC was dependent on IL-18 as pDC from IL-18-deficient mice only activated a similar proportion of NK cells as conventional DC. These data identify IL-18 as a crucial factor for pDC-mediated NK cell regulation.
Figures
References
-
- Yokoyama WM, Kim S, French AR. The dynamic life of natural killer cells. Annu. Rev. Immunol. 2004;22:405–429. - PubMed
-
- Karlhofer FM, Ribaudo RK, Yokoyama WM. MHC class I alloantigen specificity of Ly-49+ IL-2-activated natural killer cells. Nature. 1992;358:66–70. - PubMed
-
- Wagtmann N, Biassoni R, Cantoni C, Verdiani S, Malnati MS, Vitale M, Bottino C, et al. Molecular clones of the p58 NK cell receptor reveal immunoglobulin-related molecules with diversity in both the extra- and intracellular domains. Immunity. 1995;2:439–449. - PubMed
-
- Biron CA, Nguyen KB, Pien GC, Cousens LP, Salazar-Mather TP. Natural killer cells in antiviral defense: function and regulation by innate cytokines. Annu. Rev. Immunol. 1999;17:189–220. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
