

The emergence of contrast-invariant orientation tuning in simple cells of cat visual cortex
- PMID: 17408583
- PMCID: PMC1993919
- DOI: 10.1016/j.neuron.2007.02.029
The emergence of contrast-invariant orientation tuning in simple cells of cat visual cortex
Abstract
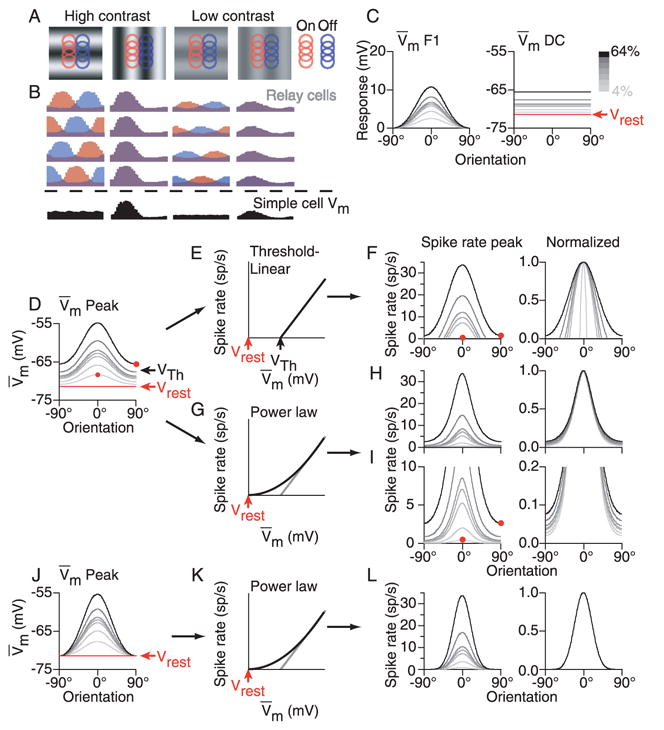
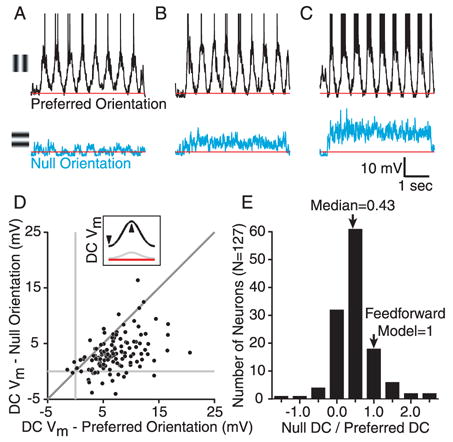
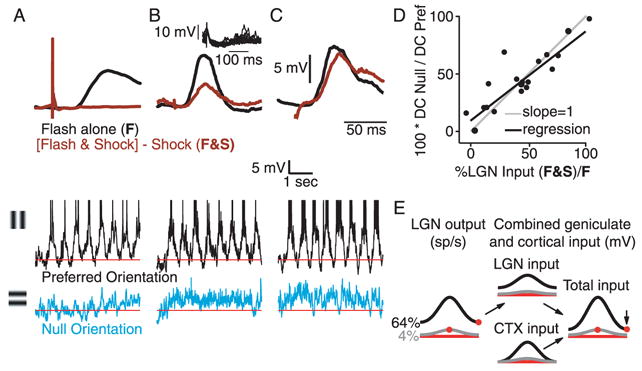
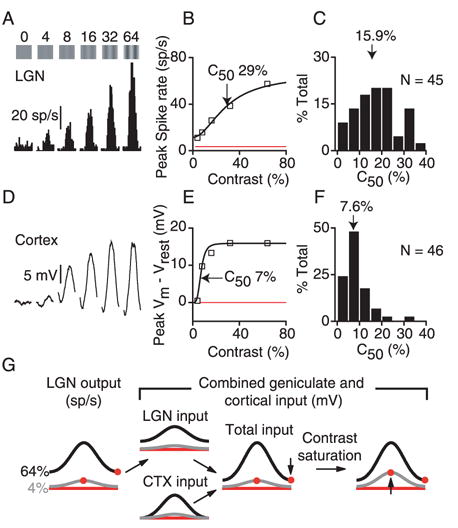
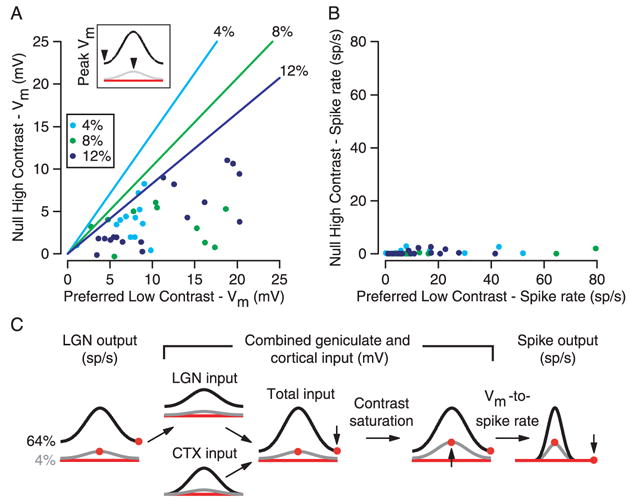
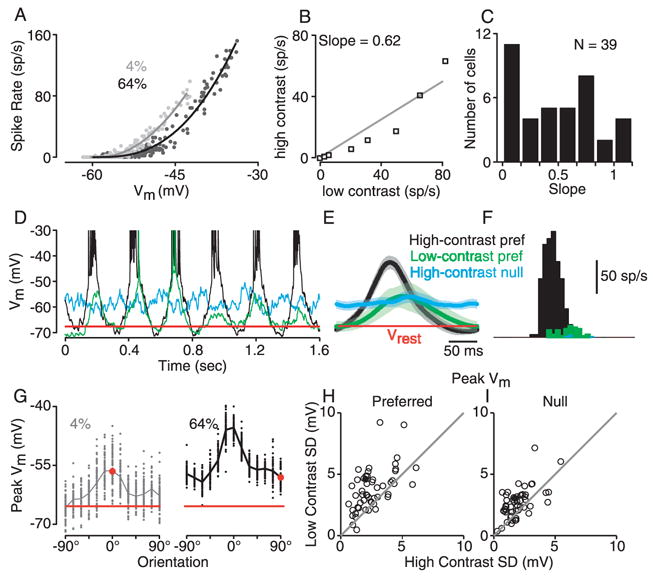
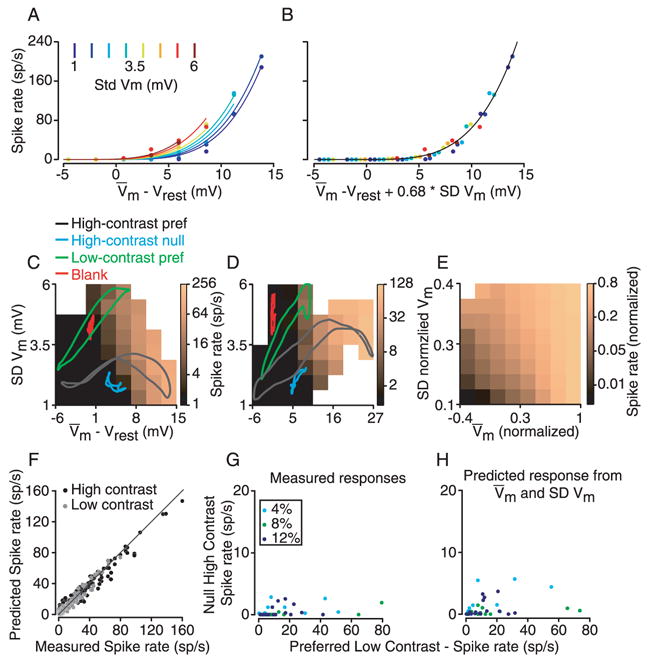
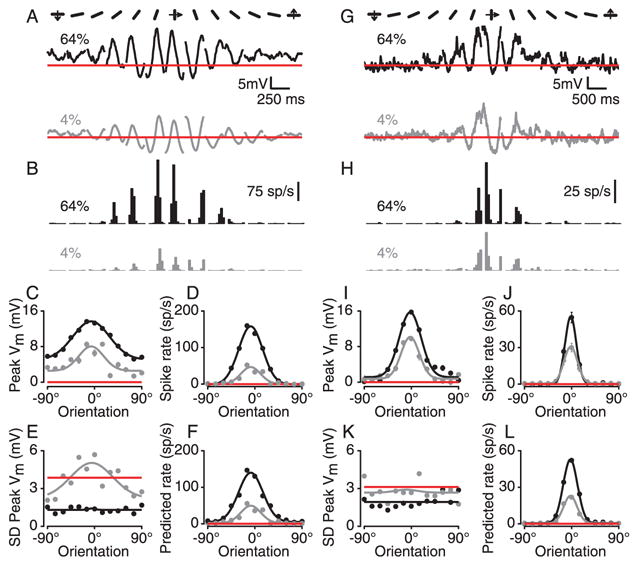
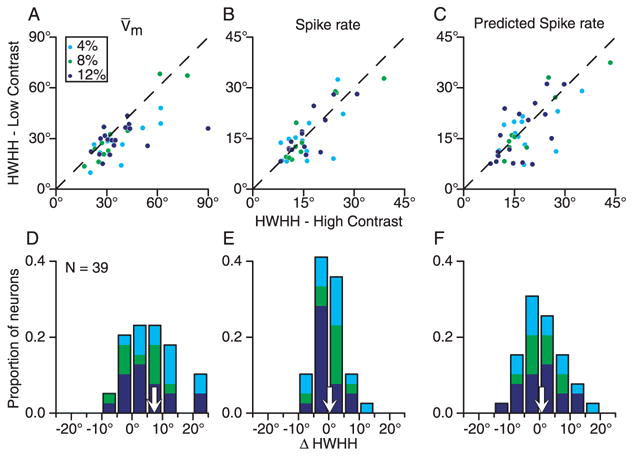
Simple cells in primary visual cortex exhibit contrast-invariant orientation tuning, in seeming contradiction to feed-forward models that rely on lateral geniculate nucleus (LGN) input alone. Contrast invariance has therefore been thought to depend on the presence of intracortical lateral inhibition. In vivo intracellular recordings instead suggest that contrast invariance can be explained by three properties of the excitatory pathway. (1) Depolarizations evoked by orthogonal stimuli are determined by the amount of excitation a cell receives from the LGN, relative to the excitation it receives from other cortical cells. (2) Depolarizations evoked by preferred stimuli saturate at lower contrasts than the spike output of LGN relay cells. (3) Visual stimuli evoke contrast-dependent changes in trial-to-trial variability, which lead to contrast-dependent changes in the relationship between membrane potential and spike rate. Thus, high-contrast, orthogonally oriented stimuli that evoke significant depolarizations evoke few spikes. Together these mechanisms, without lateral inhibition, can account for contrast-invariant stimulus selectivity.
Figures
Comment in
-
Melting the iceberg: contrast invariance in visual cortex.Neuron. 2007 Apr 5;54(1):11-3. doi: 10.1016/j.neuron.2007.03.019. Neuron. 2007. PMID: 17408573 No abstract available.
References
-
- Abbott LF, Varela JA, Sen K, Nelson SB. Synaptic depression and cortical gain control. Science. 1997;275:220–224. - PubMed
-
- Alitto HJ, Usrey WM. Influence of contrast on orientation and temporal frequency tuning in ferret primary visual cortex. J Neurophysiol. 2004;91:2797–2808. - PubMed
-
- Anderson J, Lampl I, Reichova I, Carandini M, Ferster D. Stimulus dependence of two-state fluctuations of membrane potential in cat visual cortex. Nat Neurosci. 2000a;3:617–621. - PubMed
-
- Anderson JS, Carandini M, Ferster D. Orientation tuning of input conductance, excitation, and inhibition in cat primary visual cortex. J Neurophysiol. 2000b;84:909–926. - PubMed
-
- Anderson JS, Lampl L, Gillespie D, Ferster D. The contribution of noise to contrast invariance of orientation tuning in cat visual cortex. Science. 2000c;290:1968–1971. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
