

Evolution of coadaptation in a subdivided population
- PMID: 17409093
- PMCID: PMC1893032
- DOI: 10.1534/genetics.106.063636
Evolution of coadaptation in a subdivided population
Abstract
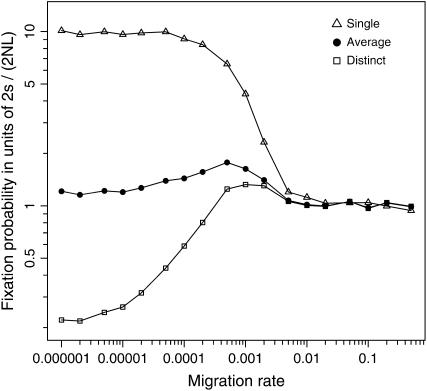
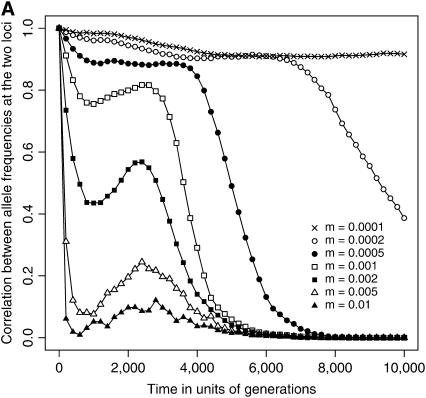
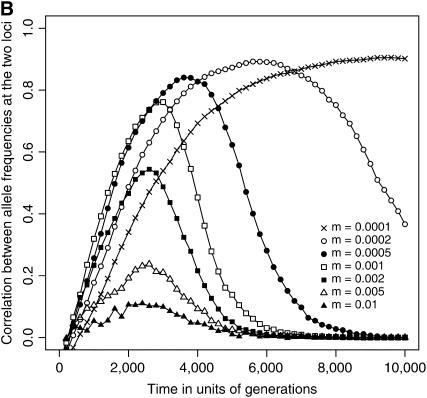
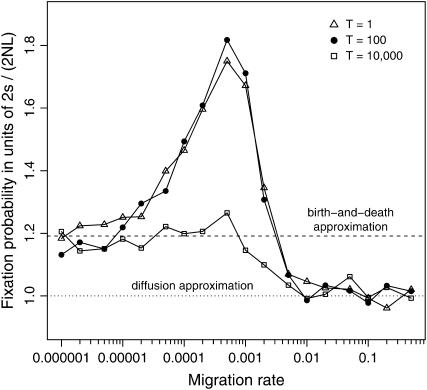
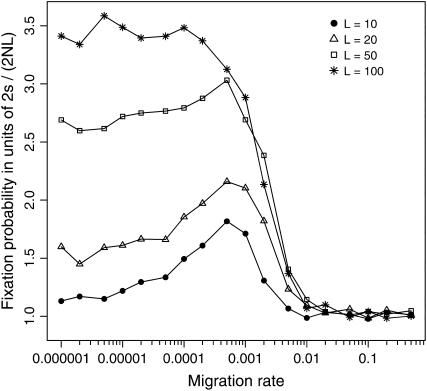
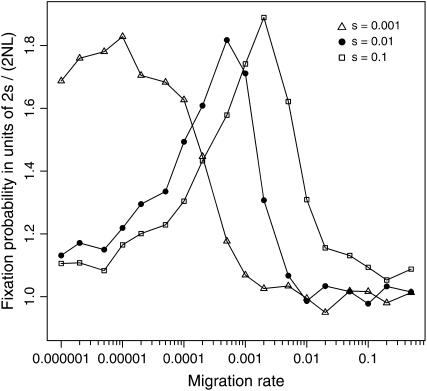
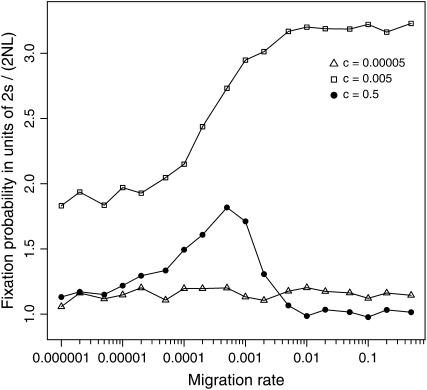
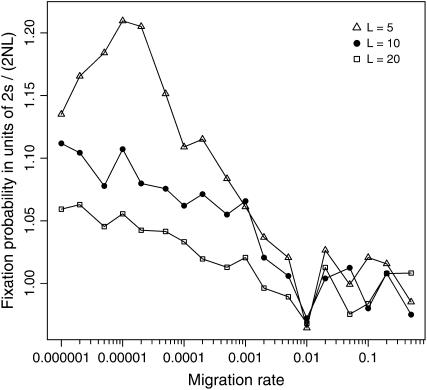
The interplay between population subdivision and epistasis is investigated by studying the fixation probability of a coadapted haplotype in a subdivided population. Analytical and simulation models are developed to study the evolutionary fate of two conditionally neutral mutations that interact epistatically to enhance fitness. We find that the fixation probability of a coadapted haplotype shows a marked increase when the population is genetically subdivided and subpopulations are loosely connected by migration. Moderate migration and isolation allow the propagation of the mutant alleles across subpopulations, while at the same time preserving the favorable allelic combination established within each subpopulation. Together they create the condition most favorable for the ultimate fixation of the coadapted haplotype. On the basis of the analytical and simulation results, we discuss the fundamental role of population subdivision and restricted gene flow in promoting the evolution of functionally integrated systems, with some implications for the shifting-balance theory of evolution.
Figures
References
-
- Barton, N. H., 1993. The probability of fixation of a favoured allele in a subdivided population. Genet. Res. 62: 149–157.
-
- Briggs, C. J., and M. F. Hoopes, 2004. Stabilizing effects in spatial parasitoid-host and predator-prey models: a review. Theor. Popul. Biol. 65: 299–315. - PubMed
-
- Chave, J., H. C. Muller-Landau and S. A. Levin, 2002. Comparing classical community models: theoretical consequences for patterns of diversity. Am. Nat. 159: 1–23. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
