

Beta-site amyloid precursor protein cleaving enzyme 1 levels become elevated in neurons around amyloid plaques: implications for Alzheimer's disease pathogenesis
- PMID: 17409228
- PMCID: PMC6672403
- DOI: 10.1523/JNEUROSCI.4396-06.2007
Beta-site amyloid precursor protein cleaving enzyme 1 levels become elevated in neurons around amyloid plaques: implications for Alzheimer's disease pathogenesis
Abstract
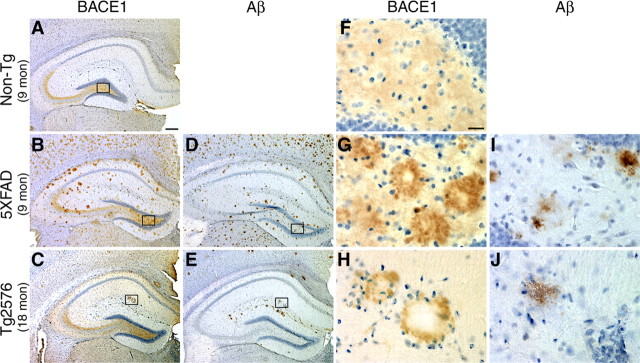
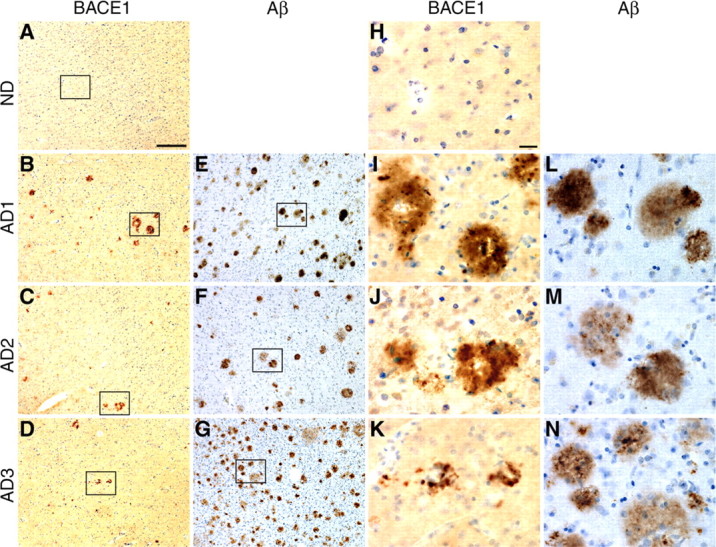
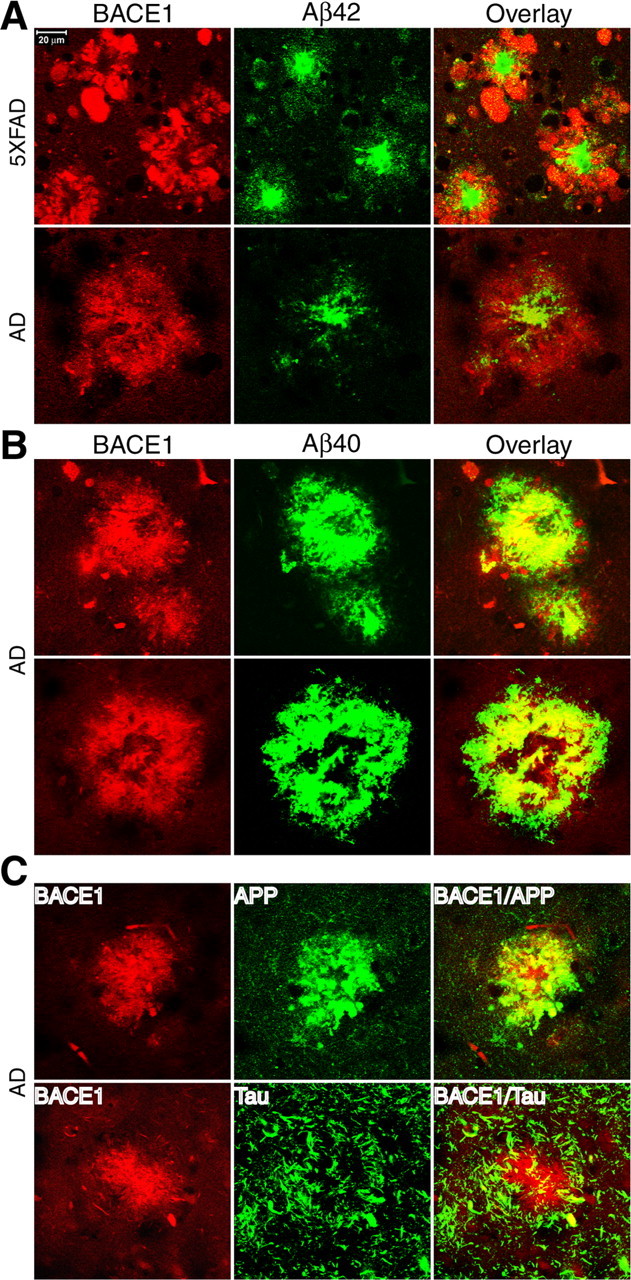
Beta-site amyloid precursor protein cleaving enzyme 1 (BACE1) (beta-secretase) initiates generation of beta-amyloid (Abeta), which plays an early role in Alzheimer's disease (AD). BACE1 levels are increased in postmortem AD brain, suggesting BACE1 elevation promotes Abeta production and AD. Alternatively, the BACE1 increase may be an epiphenomenon of late-stage AD. To distinguish between these possibilities, we analyzed BACE1 elevation using a highly specific BACE1 antibody, BACE-Cat1, made in BACE1-/- mice, which mount a robust anti-BACE1 immune response. Previous BACE1 immunohistochemical studies lack consistent results because typical BACE1 antibodies produce nonspecific background, but BACE-Cat1 immunolabels BACE1 only. BACE1 elevation was recapitulated in two amyloid precursor protein (APP) transgenic mouse lines. 5XFAD mice form amyloid plaques at young ages and exhibit neuron loss. In contrast, Tg2576 form plaques at a more advanced age and do not show cell death. These two mouse lines allow differentiation between early Abeta-induced events and late phenomena related to neuron death. BACE1 levels became elevated in parallel with amyloid burden in each APP transgenic, starting early in 5XFAD and late in Tg2576. The increase in BACE1 protein occurred without any change in BACE1 mRNA level, indicating a posttranscriptional mechanism. In APP transgenic and AD brains, high BACE1 levels were observed in an annulus around Abeta42-positive plaque cores and colocalized with neuronal proteins. These results demonstrate that amyloid plaques induce BACE1 in surrounding neurons at early stages of pathology before neuron death occurs. We conclude that BACE1 elevation is most likely triggered by the amyloid pathway and may drive a positive-feedback loop in AD.
Figures



References
-
- Apelt J, Bigl M, Wunderlich P, Schliebs R. Aging-related increase in oxidative stress correlates with developmental pattern of beta-secretase activity and beta-amyloid plaque formation in transgenic Tg2576 mice with Alzheimer-like pathology. Int J Dev Neurosci. 2004;22:475–484. - PubMed
-
- Blasko I, Beer R, Bigl M, Apelt J, Franz G, Rudzki D, Ransmayr G, Kampfl A, Schliebs R. Experimental traumatic brain injury in rats stimulates the expression, production and activity of Alzheimer's disease beta-secretase (BACE-1) J Neural Transm. 2004;111:523–536. - PubMed
-
- Bodendorf U, Danner S, Fischer F, Stefani M, Sturchler-Pierrat C, Wiederhold KH, Staufenbiel M, Paganetti P. Expression of human beta-secretase in the mouse brain increases the steady-state level of beta-amyloid. J Neurochem. 2002;80:799–806. - PubMed
-
- Cai H, Wang Y, McCarthy D, Wen H, Borchelt DR, Price DL, Wong PC. BACE1 is the major beta-secretase for generation of Abeta peptides by neurons. Nat Neurosci. 2001;4:233–234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous