

doi: 10.1126/science.1141314.
Epub 2007 Apr 5.
Genes required for mitotic spindle assembly in Drosophila S2 cells
Affiliations
- PMID: 17412918
- PMCID: PMC2837481
- DOI: 10.1126/science.1141314
Item in Clipboard
Genes required for mitotic spindle assembly in Drosophila S2 cells
Science.
.
Abstract
The formation of a metaphase spindle, a bipolar microtubule array with centrally aligned chromosomes, is a prerequisite for the faithful segregation of a cell's genetic material. Using a full-genome RNA interference screen of Drosophila S2 cells, we identified about 200 genes that contribute to spindle assembly, more than half of which were unexpected. The screen, in combination with a variety of secondary assays, led to new insights into how spindle microtubules are generated; how centrosomes are positioned; and how centrioles, centrosomes, and kinetochores are assembled.
Figures
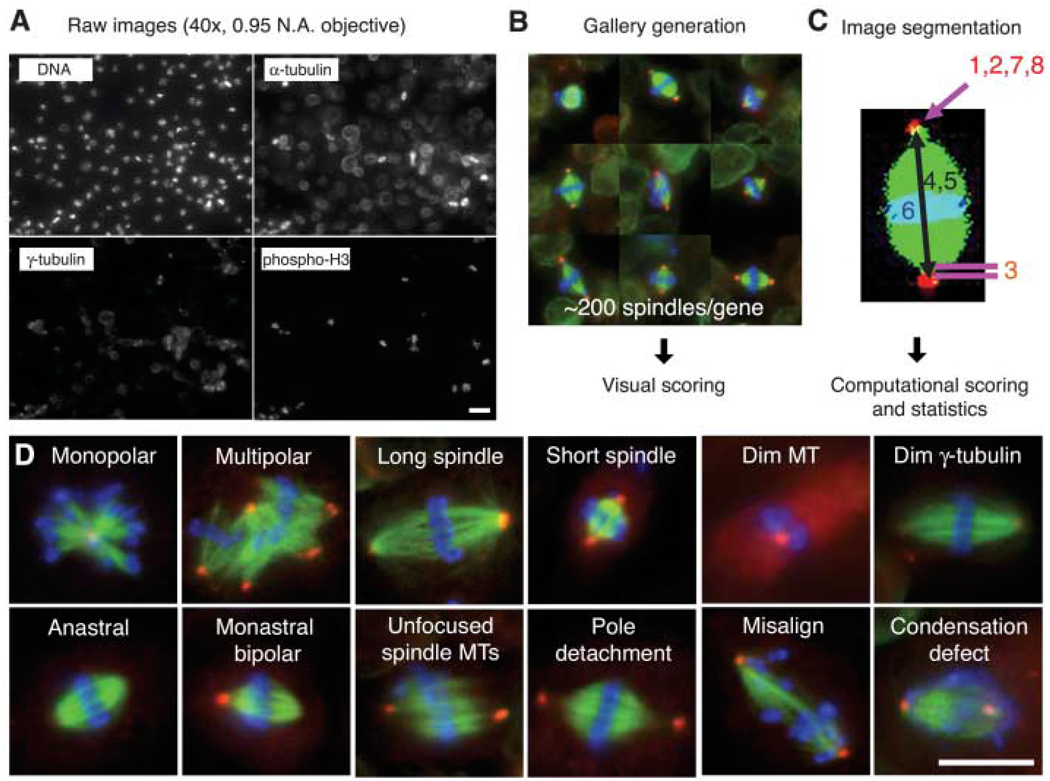
An image-based, genome-wide RNAi screening ofmetaphase spindlemorphology. (A) Drosophila S2 cells were treated with dsRNAs of 14,425 genes, along with Cdc27/Apc3 dsRNA, to accumulate metaphase cells. After 4 days, cells were transferred and adhered onto ConA-coated glass-bottom plates. Cells were immunostained for DNA, α-tubulin, γ-tubulin and pH3 and imaged by high-throughput automated microscopy. Scale bar, 20 µm. (B) Mitotic cells were automatically selected by a computer algorithm that detects pH3 staining. The selected ~200 mitotic cells (mostly metaphase) were displayed in a gallery for an observer to score phenotypes. (C) Phenotypes were also analyzed by computer after image segmentation (1, monopolar; 2, multipolar; 3, pole detachment; 4, long spindle; 5, short spindle; 6, misalignment; 7, large γ-tubulin area; 8, dim γ-tubulin) (11). (D) Twelve major phenotypes identified in the screen. Scale bar, 10 µm.
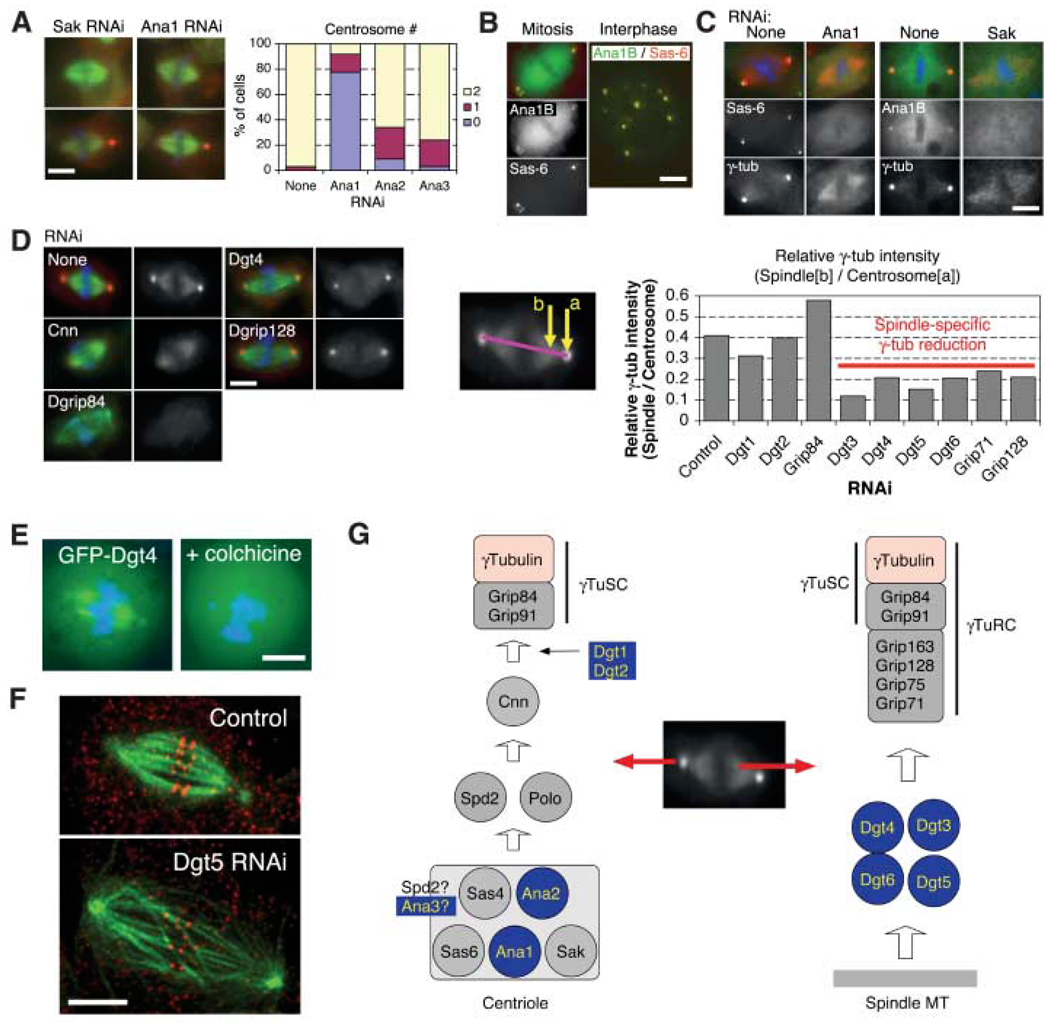
Genes required for localizing γ-tubulin to the centrosome and the spindle. (A) (Left) Anastral as well as monastral bipolar spindles were observed after RNAi to Sak or three previously unknown Ana genes (the latter two shown in fig. S3). (Right) Centrosome number was counted for bipolar spindles after four rounds of RNAi treatment. (B) Ana1 (isoform B) was colocalized with the centriole marker mRFP-Sas-6. (C) Ana1 RNAi caused the loss of GFP-Sas-6 from the pole region of anastral spindles, whereas SAK RNAi interfered with GFP-Ana1B localization to poles. (D) (Left) Various dim γ-tubulin RNAi phenotypes. The γ-tubulin signal is reduced at the centrosome alone (Cnn), at both the centrosome and spindle (Dgrip84), or only at the spindle (Dgt4 and Dgrip128). (Right) Intensity at centrosome (a) and spindle region (b) was measured, and the relative intensity was calculated (11). (E) Spindle localization of GFP-Dgt4 is lost after colchicine-induced MT depolymerization. (F) RNAi of Dgt5 (as well as other genes required for spindle localization of γ-tubulin) produces long spindles with low MT density and misaligned chromosomes. Shown is a maximum intensity projection of four optical slices obtained at 0.25-µm intervals by spinning-disk confocal microscope. Red, antibody to CENP-A; green, MT. Scale bar, 5 µm. (G) Model for recruitment pathway of γ-tubulin to the centrosome and spindle MTs. Previously unknown or unexpected genes are highlighted in blue in this and other figures.
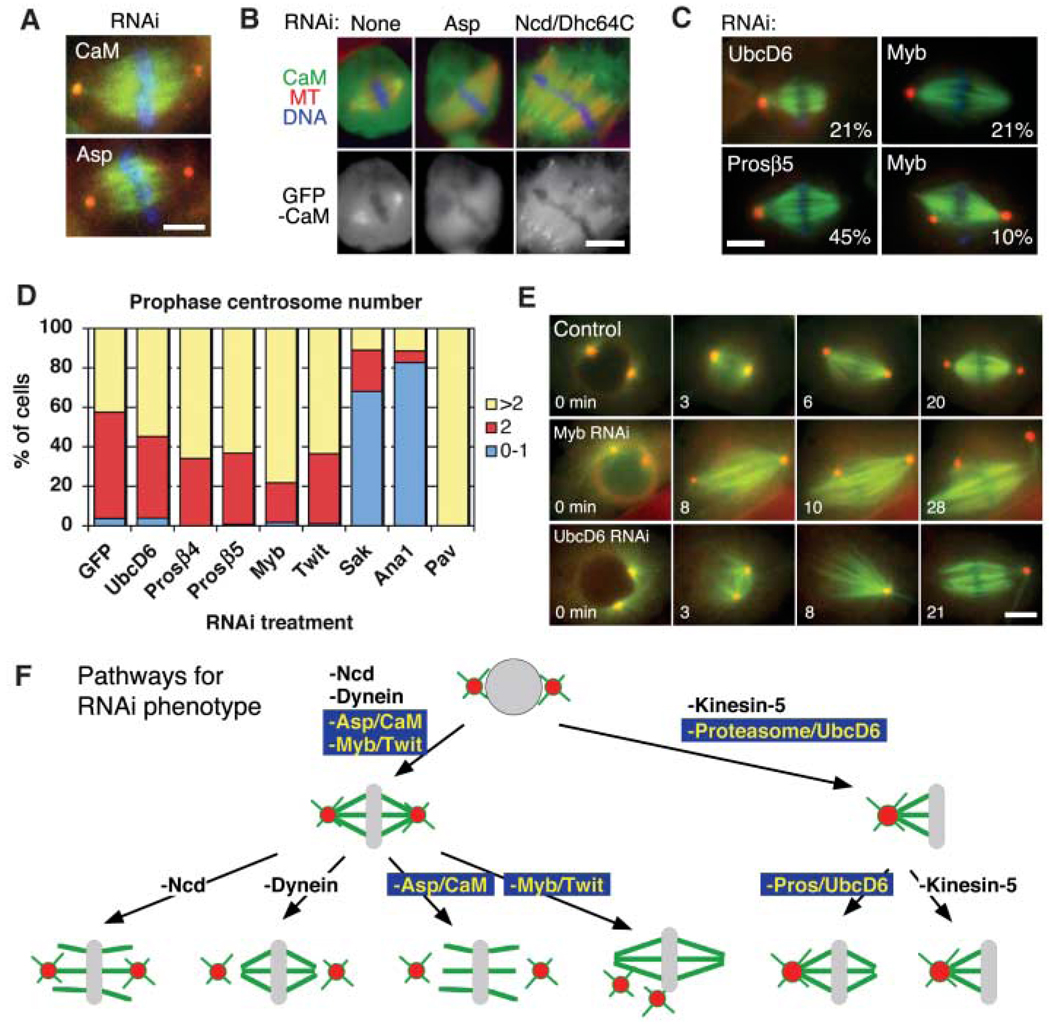
Genes required for centrosome separation and association to spindle poles. (A) Identical kinetochore-fiber unfocusing and centrosome-detachment phenotypes were observed after Asp and calmodulin (CaM) RNAi. See movie S2. (B) GFP-CaM was localized at spindle poles in an Asp-dependent manner, but was still detected at unfocused poles induced by Ncd/Dhc64C RNAi. (C) Monastral bipolar spindles produced by UbcD6, proteasome subunit β5, or Myb RNAi (percentage of cells with phenotype indicated; 3% to 8% for untreated cells). Centrosome detachment also was detected after Myb RNAi. (D) At prophase, most proteosome (Prosβ4 and Prosβ5), UbcD6, Myb, and Twit RNAi cells (n > 65) had >1 centrosome; Sak, Ana1 (centriole duplication), and Pav (cytokinesis) served as control RNAis that decrease or increase centrosome numbers. (E) Time-lapse imaging of γ-tubulin-GFP (red) and mCherry-tubulin (green) from prophase to metaphase shows bipolar spindle formation, and then centrosome detachment after Myb RNAi (80%) (n = 20), and immediate centrosome fusion after NEB for UbcD6 RNAi (25%) (n = 20). See movie S3. Scale bars, 5 µm. (F) Models for various spindle-pole phenotypes in response to various protein depletions.
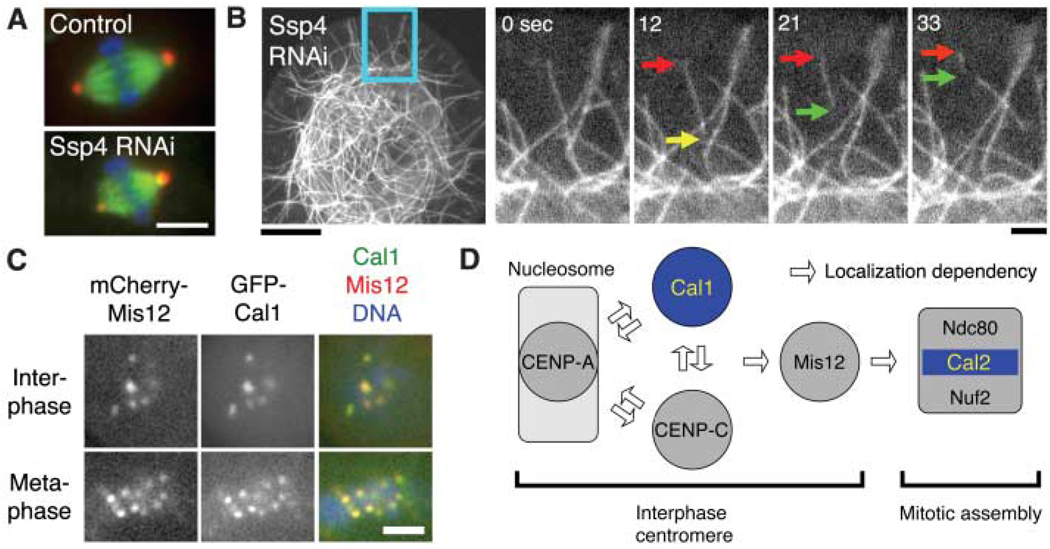
Regulation of spindle length and chromosome alignment. (A) Spindle length was altered after RNAi depletion of the novel protein Ssp4. Scale bar, 5 µm. (B) MT severing (yellow arrow) frequently occurred after Ssp4 RNAi. Severed MTs often showed treadmilling behavior (red and green arrows) and then disappeared. Scale bars, 10 µm (left), 2 µm (right). See also movie S4. (C) Previously unknown Cal1 protein localizes to the centromere (marked by mCherry-Mis12). (Localization data for other proteins are in fig. S7). Scale bar, 2 µm. (D) Model for kinetochore assembly in S2 cells based on protein localization and RNAi. (Data are in fig. S7, D to F).
References
-
- Mitchison TJ, Salmon ED. Nat. Cell Biol. 2001;3:E17. - PubMed
-
- McIntosh JR, Grishchuk EL, West RR. Annu. Rev. Cell Dev. Biol. 2002;18:193. - PubMed
-
- Rieder CL, Khodjakov A. Science. 2003;300:91. - PubMed
-
- Gadde S, Heald R. Curr. Biol. 2004;14:R797. - PubMed
-
- Scholey JM, Brust-Mascher I, Mogilner A. Nature. 2003;422:746. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
