

Mice lacking inhibitory leptin receptor signals are lean with normal endocrine function
- PMID: 17415414
- PMCID: PMC1838925
- DOI: 10.1172/JCI30688
Mice lacking inhibitory leptin receptor signals are lean with normal endocrine function
Abstract
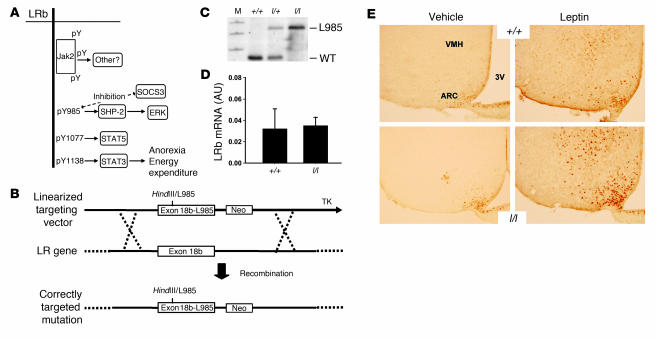
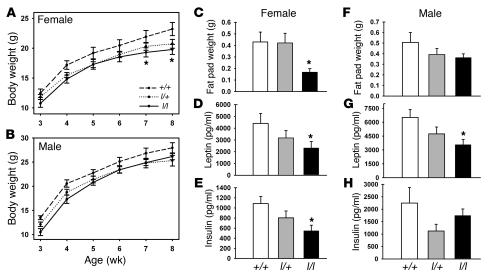
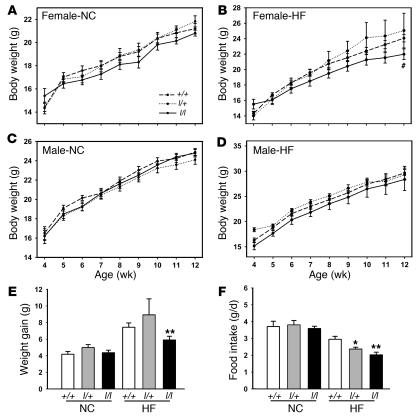
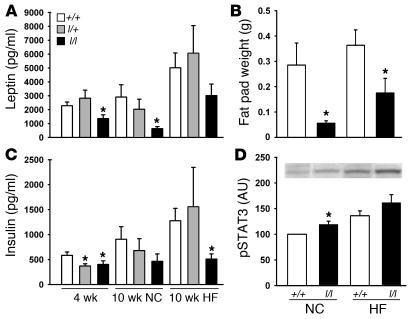
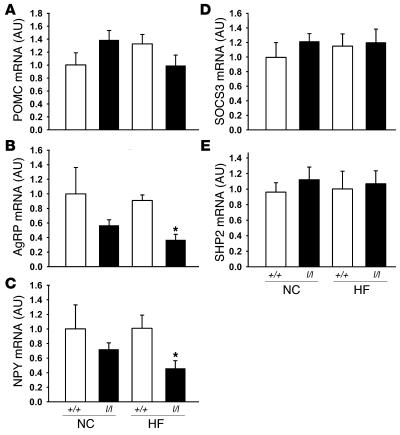
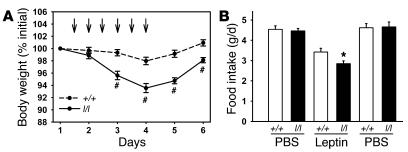
The adipose-derived hormone, leptin, acts via its receptor (LRb) to convey the status of body energy stores to the brain, decreasing feeding and potentiating neuroendocrine energy expenditure. The failure of high levels of leptin in most obese individuals to promote weight loss defines a state of diminished responsiveness to increased leptin, termed leptin resistance. Leptin stimulates the phosphorylation of several tyrosine residues on LRb to mediate leptin action. We homologously replaced LRb in mice with a receptor with a mutation in one of these sites (Tyr985) in order to examine its role in leptin action and signal attenuation in vivo. Mice homozygous for this mutation are neuroendocrinologically normal, but females demonstrate decreased feeding, decreased expression of orexigenic neuropeptides, protection from high-fat diet-induced obesity, and increased leptin sensitivity in a sex-biased manner. Thus, leptin activates autoinhibitory signals via LRb Tyr985 to attenuate the anti-adiposity effects of leptin, especially in females, potentially contributing to leptin insensitivity in obesity.
Figures
References
-
- Myers M.G., Jr. Leptin receptor signaling and the regulation of mammalian physiology. Recent Prog. Horm. Res. 2004;59:287–304. - PubMed
-
- Elmquist J.K., Coppari R., Balthasar N., Ichinose M., Lowell B.B. Identifying hypothalamic pathways controlling food intake, body weight, and glucose homeostasis. J. Comp. Neurol. 2005;493:63–71. - PubMed
-
- Ahima R.S., et al. Role of leptin in the neuroendocrine response to fasting. Nature. 1996;382:250–252. - PubMed
-
- Zhang Y., et al. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994;372:425–432. - PubMed
-
- Chua S.C., Jr. et al. Phenotypes of mouse diabetes and rat fatty due to mutations in the OB (Leptin) receptor. . Science. 1996;271:994–996. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
