

Thrombospondin-1 inhibits nitric oxide signaling via CD36 by inhibiting myristic acid uptake
- PMID: 17416590
- PMCID: PMC2430148
- DOI: 10.1074/jbc.M701638200
Thrombospondin-1 inhibits nitric oxide signaling via CD36 by inhibiting myristic acid uptake
Abstract
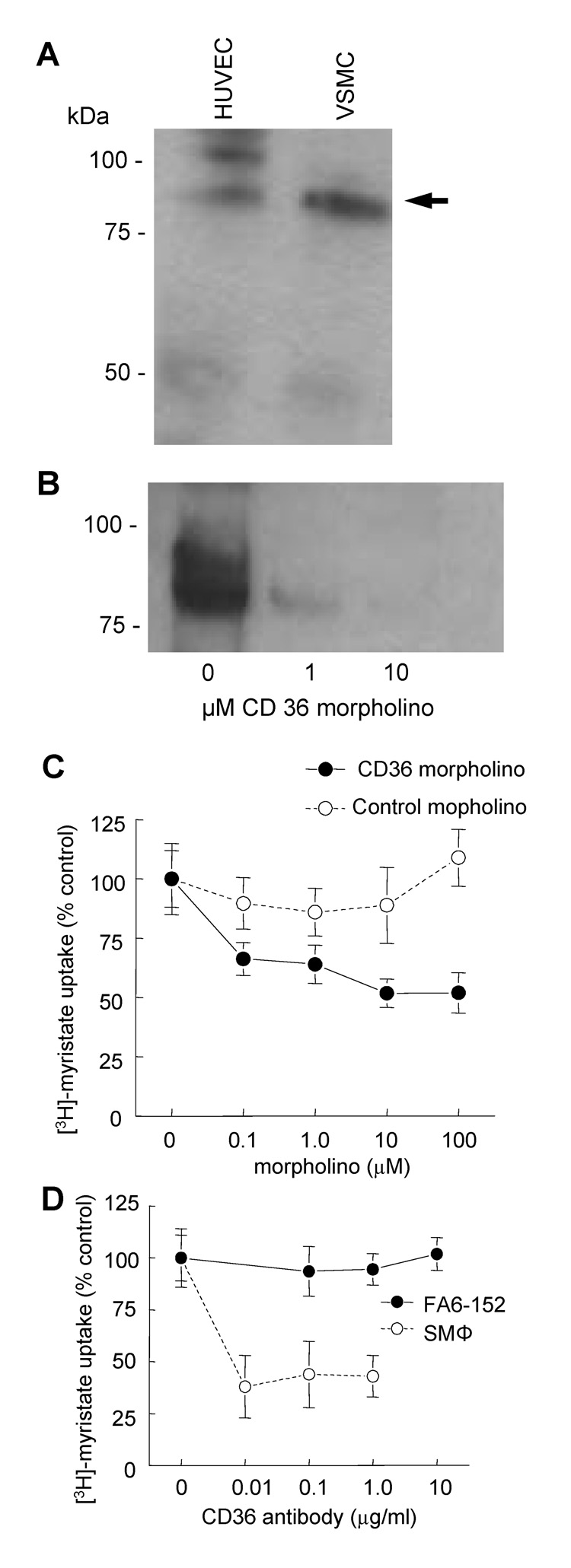
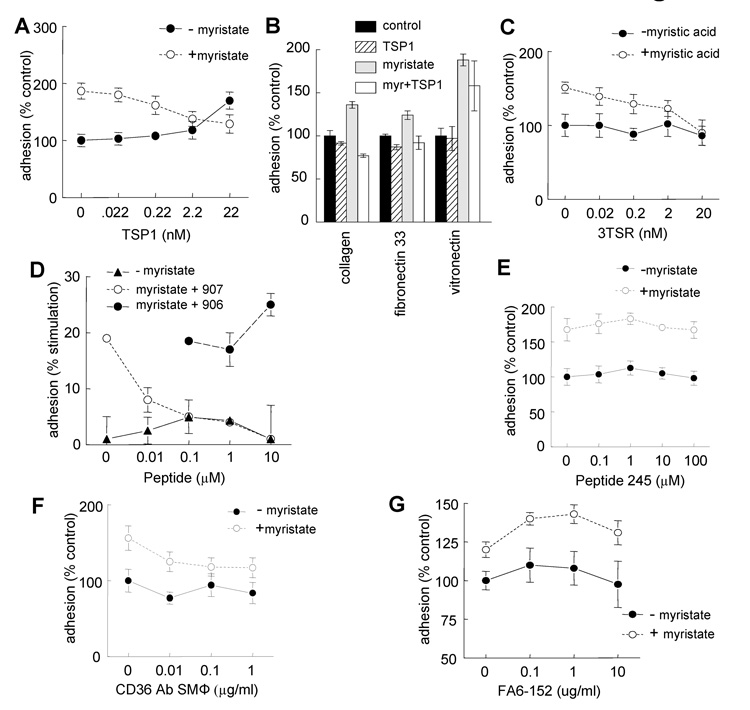
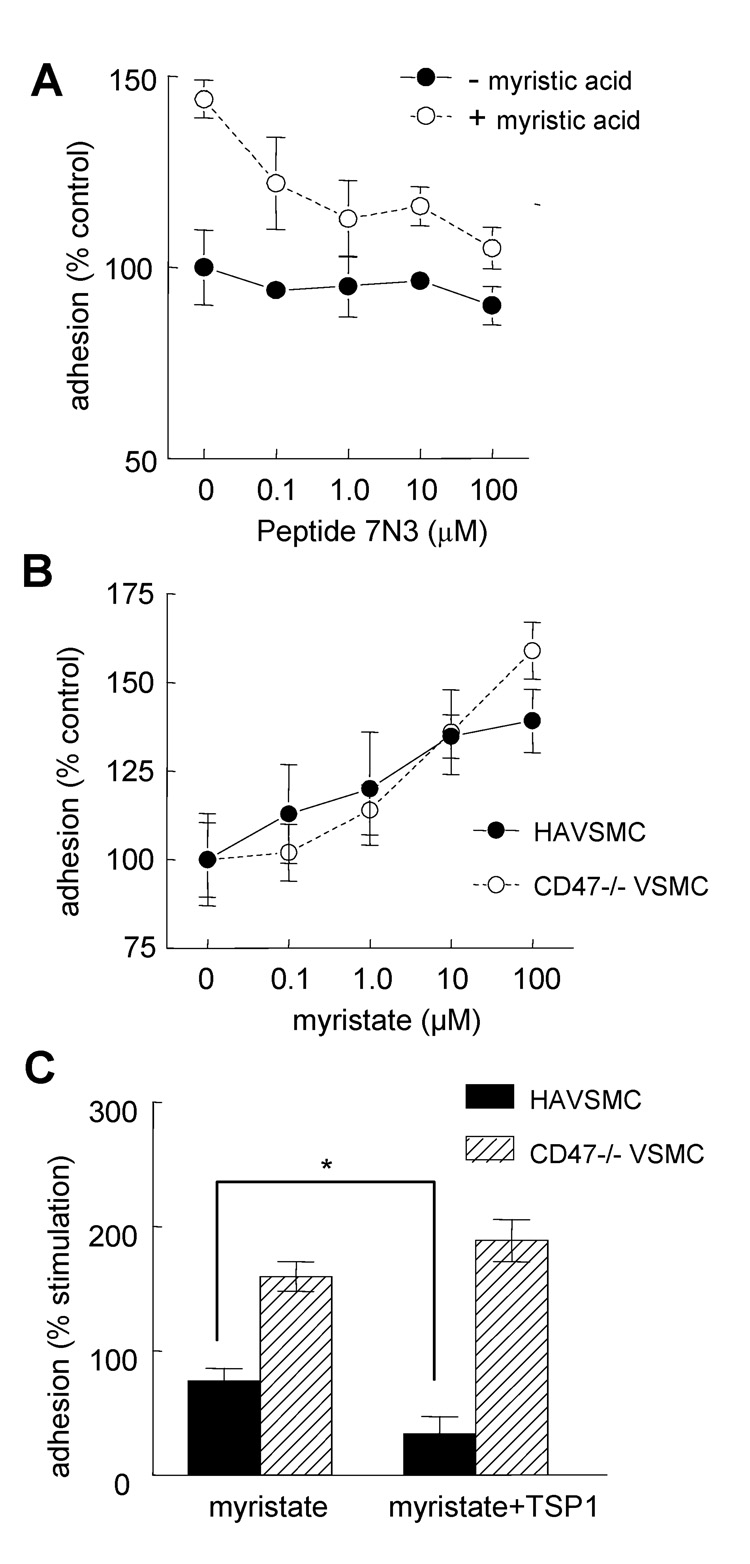
Although CD36 is generally recognized to be an inhibitory signaling receptor for thrombospondin-1 (TSP1), the molecular mechanism for transduction of this signal remains unclear. Based on evidence that myristic acid and TSP1 each modulate endothelial cell nitric oxide signaling in a CD36-dependent manner, we examined the ability of TSP1 to modulate the fatty acid translocase activity of CD36. TSP1 and a CD36 antibody that mimics the activity of TSP1 inhibited myristate uptake. Recombinant TSP1 type 1 repeats were weakly inhibitory, but an anti-angiogenic peptide derived from this domain potently inhibited myristate uptake. This peptide also inhibited membrane translocation of the myristoylated CD36 signaling target Fyn and activation of Src family kinases. Myristate uptake stimulated cGMP synthesis via endothelial nitric-oxide synthase and soluble guanylyl cyclase. CD36 ligands blocked myristate-stimulated cGMP accumulation in proportion to their ability to inhibit myristate uptake. TSP1 also inhibited myristate-stimulated cGMP synthesis by engaging its receptor CD47. Myristate stimulated endothelial and vascular smooth muscle cell adhesion on type I collagen via the NO/cGMP pathway, and CD36 ligands that inhibit myristate uptake blocked this response. Therefore, the fatty acid translocase activity of CD36 elicits proangiogenic signaling in vascular cells, and TSP1 inhibits this response by simultaneously inhibiting fatty acid uptake via CD36 and downstream cGMP signaling via CD47.
Figures






References
-
- Carmeliet P. Nature. 2005;438(7070):932–936. - PubMed
-
- Lawler J, Detmar M. Int J Biochem Cell Biol. 2004;36(6):1038–1045. - PubMed
-
- Nyberg P, Xie L, Kalluri R. Cancer Res. 2005;65(10):3967–3979. - PubMed
-
- Calzada MJ, Zhou L, Sipes JM, Zhang J, Krutzsch HC, Iruela-Arispe ML, Annis DS, Mosher DF, Roberts DD. Circ Res. 2004;94(4):462–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
