

Beta-arrestins 1 and 2 differentially regulate LPS-induced signaling and pro-inflammatory gene expression
- PMID: 17418896
- PMCID: PMC1945129
- DOI: 10.1016/j.molimm.2007.02.009
Beta-arrestins 1 and 2 differentially regulate LPS-induced signaling and pro-inflammatory gene expression
Abstract
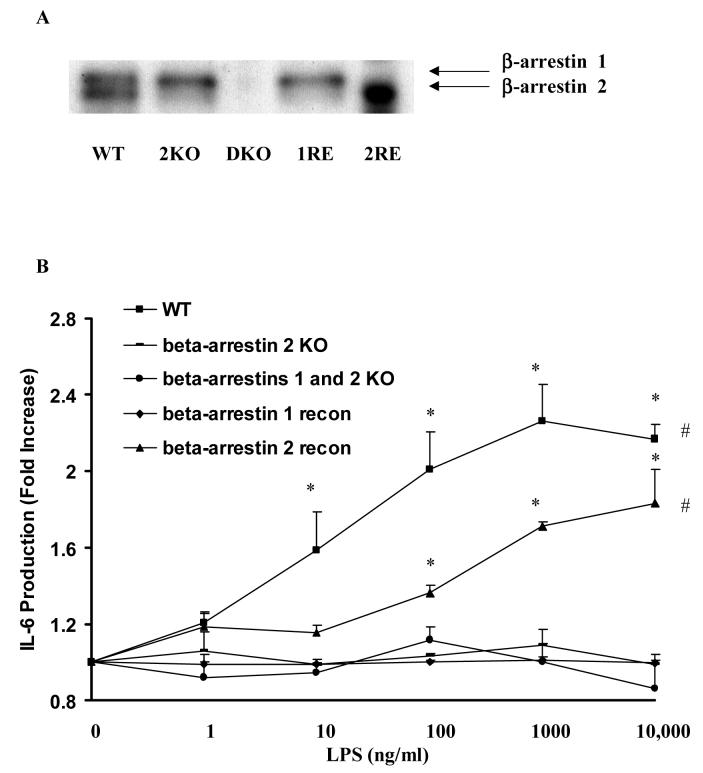
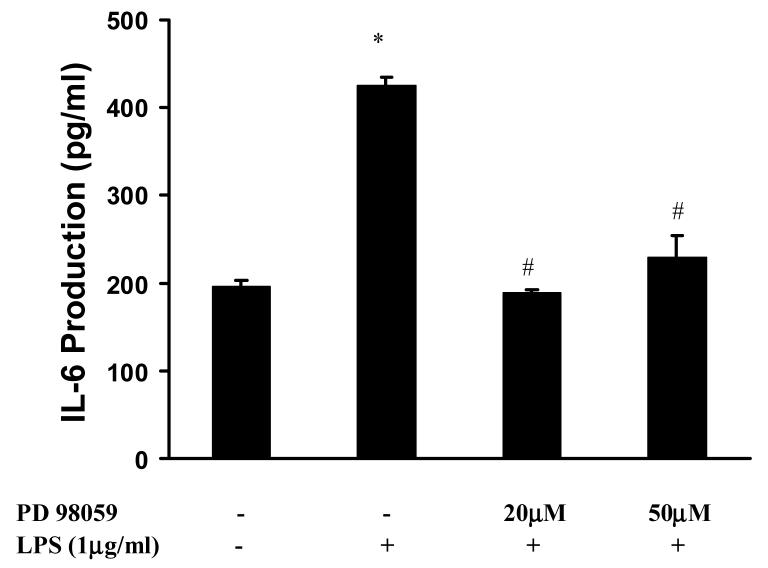
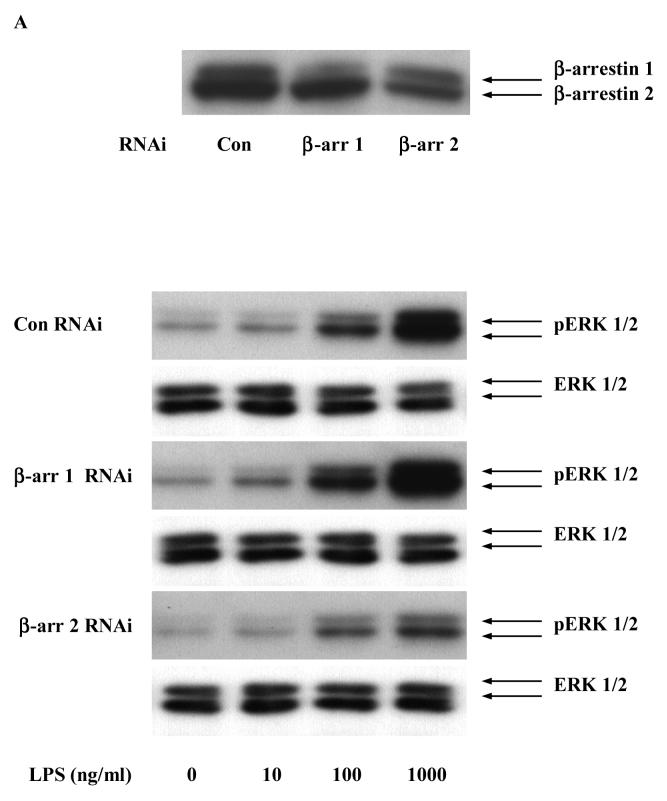
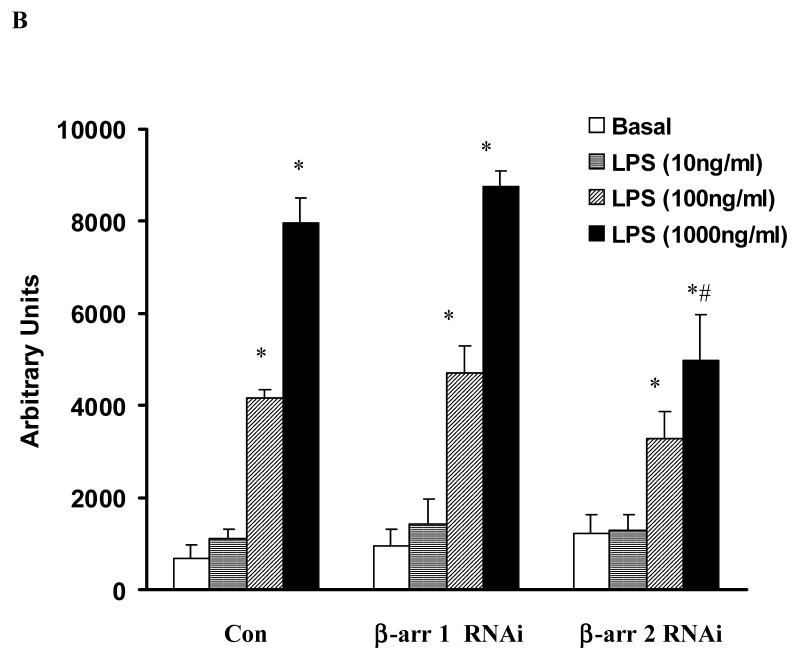
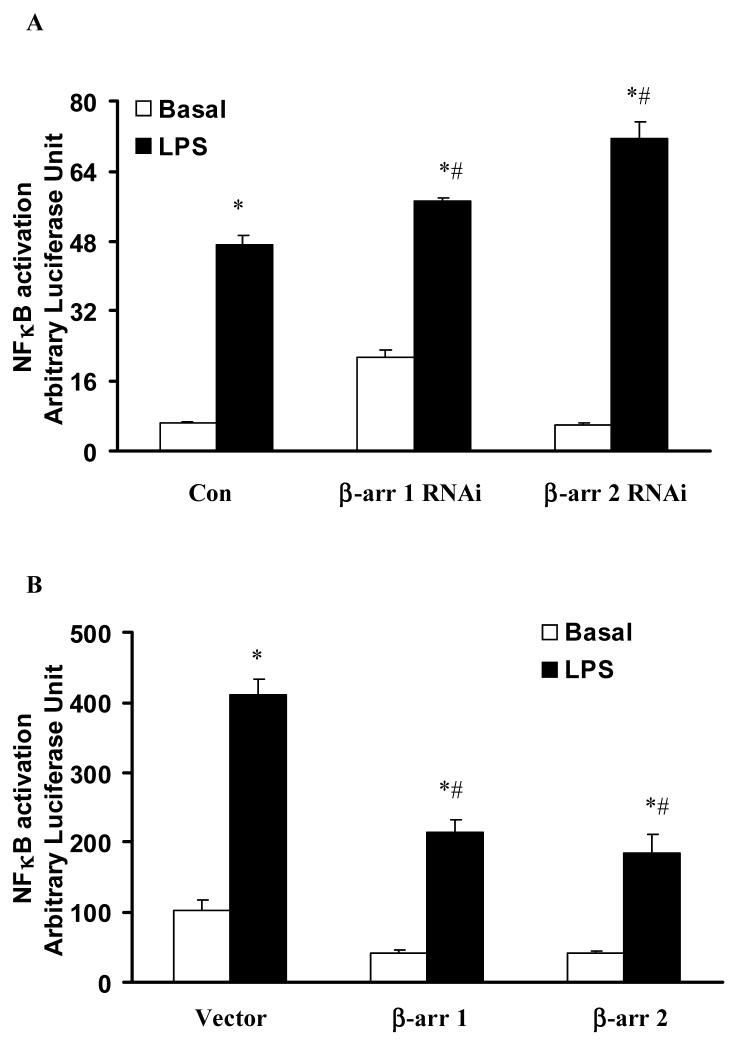
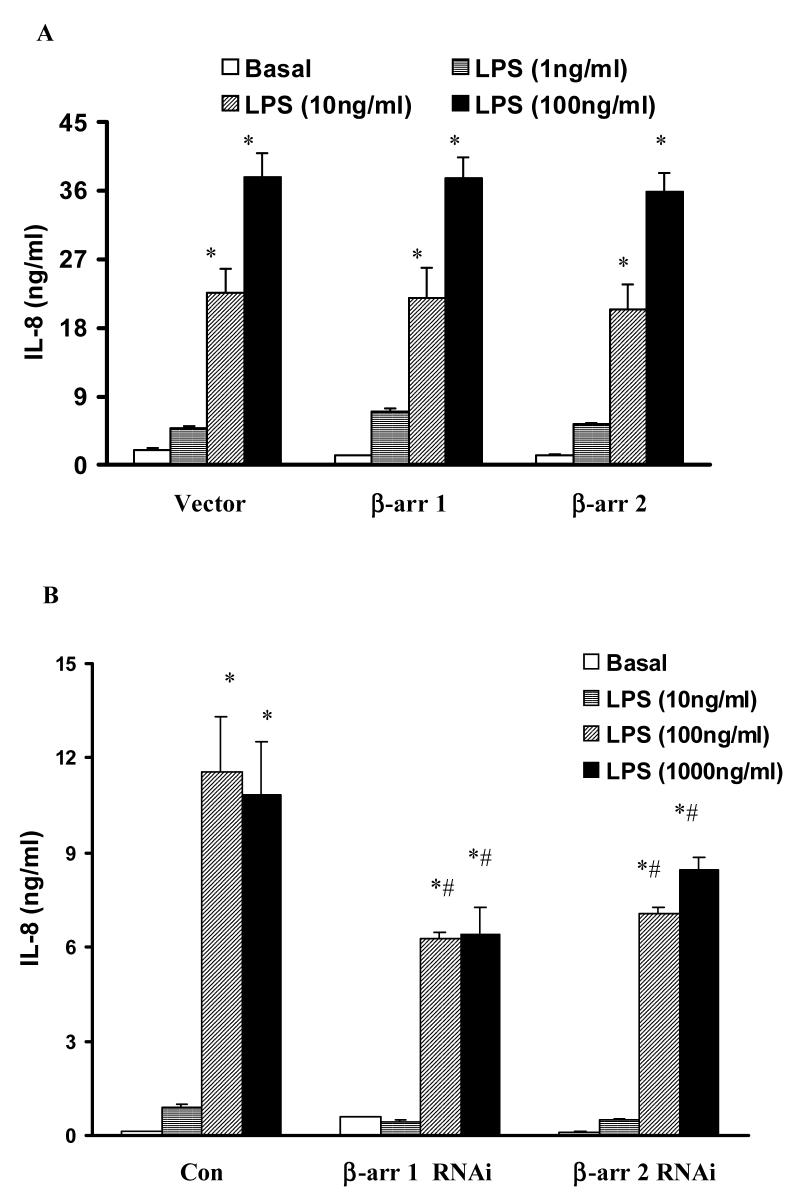
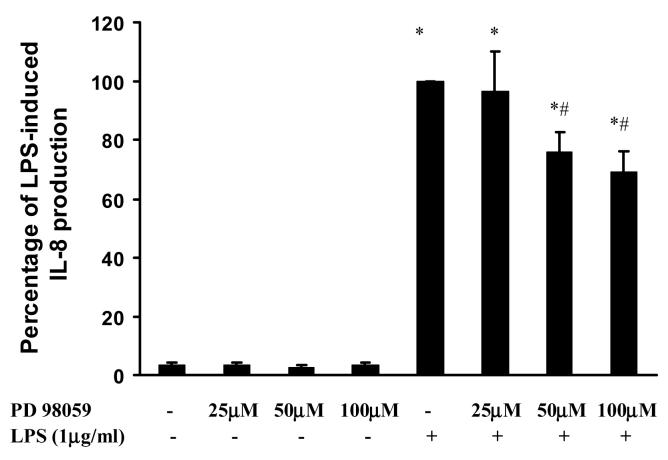
Toll like receptors, the critical receptor family in innate immunity, have been shown to signal via both ERK 1/2 and transcription factor NFkappaB. beta-Arrestins 1 and 2 have recently been implicated in modulation of NFkappaB signaling and ERK 1/2 activation. Using a number of approaches: mouse embryonic fibroblasts (MEF) from wild-type (WT), beta-arrestins knockouts (KO), beta-arrestins 1 and 2 double KO, and MEFs with reconstituted WT beta-arrestins in the double KO cells, RNA interference (siRNA) specific knockdown of beta-arrestins, and overexpression of WT beta-arrestins, it was demonstrated that beta-arrestin 2 positively regulates LPS-induced ERK 1/2 activation and both beta-arrestins 1 and 2 negatively regulate LPS-induced NFkappaB activation. Also beta-arrestin 2 positively regulate LPS-induced IL-6 production and both beta-arrestins 1 and 2 positively regulate LPS-induced IL-8 production. The specific ERK1/2 inhibitor PD98059 significantly decreased LPS-induced IL-6 and IL-8 production suggesting that IL-6 and IL-8 production is, in part, mediated by ERK 1/2 activation. Over expression of wild type beta-arrestins 1 and 2 had no effect on LPS-induced ERK1/2 activation and LPS-induced IL-8 production suggesting that endogenous beta-arrestins 1 and 2 are sufficient to mediate maximum ERK 1/2 activity and IL-8 production. beta-Arrestins thus not only negatively regulate LPS-induced NFkappaB activation but also positively regulate ERK 1/2 activation and specific pro-inflammatory gene expression. Understanding the role of beta-arrestins in regulation of TLR signaling pathways may provide novel insights into control mechanisms for inflammatory gene expression.
Figures
References
-
- Ahn S, Wei H, Garrison TR, Lefkowitz RJ. Reciprocal regulation of angiotensin receptor-activated extracellular signal-regulated kinases by beta-arrestins 1 and 2. J. Biol. Chem. 2004;279:7807–7811. - PubMed
-
- Akashi S, Shimazu R, Ogata H, Nagai Y, Takeda K, Kimoto M, Miyake K. Cutting edge: cell surface expression and lipopolysaccharide signaling via the toll-like receptor 4-MD-2 complex on mouse peritoneal macrophages. J. Immunol. 2000;164:3471–3475. - PubMed
-
- Akira S, Takeda K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004;4:499–511. - PubMed
-
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
