

Cortical mechanisms of action selection: the affordance competition hypothesis
- PMID: 17428779
- PMCID: PMC2440773
- DOI: 10.1098/rstb.2007.2054
Cortical mechanisms of action selection: the affordance competition hypothesis
Abstract
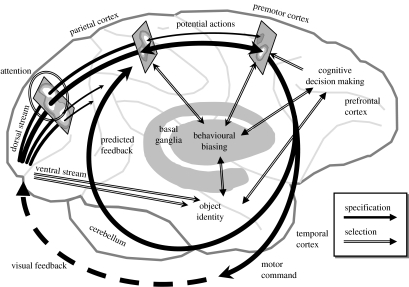
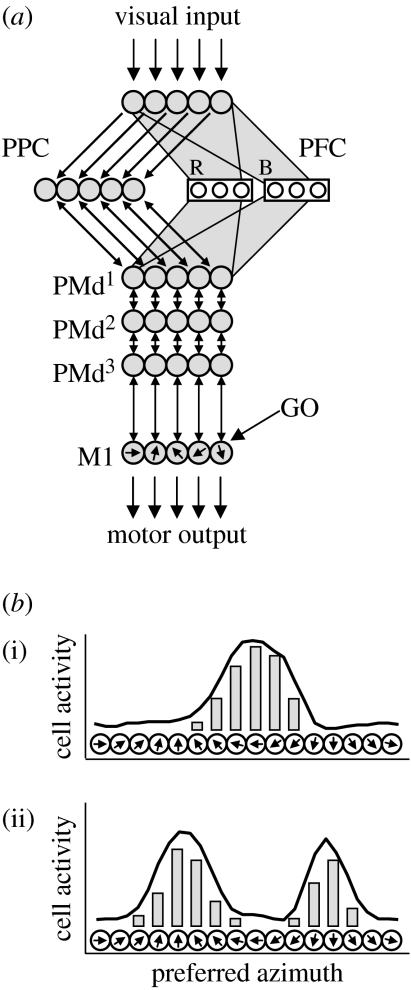
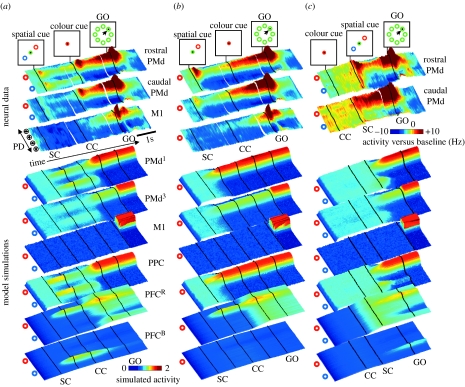
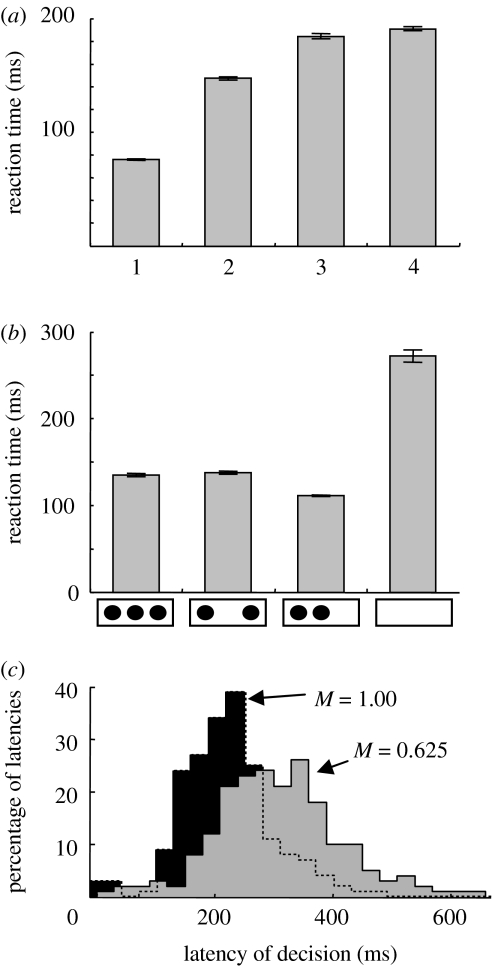
At every moment, the natural world presents animals with two fundamental pragmatic problems: selection between actions that are currently possible and specification of the parameters or metrics of those actions. It is commonly suggested that the brain addresses these by first constructing representations of the world on which to build knowledge and make a decision, and then by computing and executing an action plan. However, neurophysiological data argue against this serial viewpoint. In contrast, it is proposed here that the brain processes sensory information to specify, in parallel, several potential actions that are currently available. These potential actions compete against each other for further processing, while information is collected to bias this competition until a single response is selected. The hypothesis suggests that the dorsal visual system specifies actions which compete against each other within the fronto-parietal cortex, while a variety of biasing influences are provided by prefrontal regions and the basal ganglia. A computational model is described, which illustrates how this competition may take place in the cerebral cortex. Simulations of the model capture qualitative features of neurophysiological data and reproduce various behavioural phenomena.
Figures
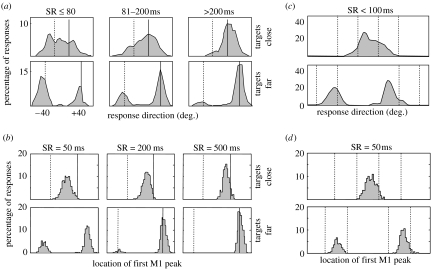
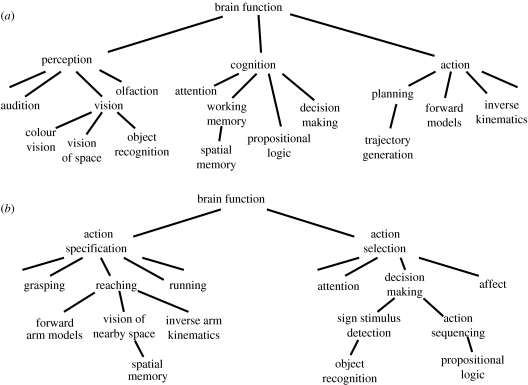
References
-
- Adams F, Mele A. The role of intention in intentional action. Can. J. Philos. 1989;19:511–531.
-
- Alexander G.E, Crutcher M.D. Functional architecture of basal ganglia circuits: neural substrates of parallel processing. Trends Neurocogn. Sci. 1990a;13:266–271. doi:10.1016/0166-2236(90)90107-L - DOI - PubMed
-
- Alexander G.E, Crutcher M.D. Neural representations of the target (goal) of visually guided arm movements in three motor areas of the monkey. J. Neurophysiol. 1990b;64:164–178. - PubMed
-
- Alexander G.E, Crutcher M.D. Preparation for movement: neural representations of intended direction in three motor areas of the monkey. J. Neurophysiol. 1990c;64:133–150. - PubMed
-
- Allport D.A. Selection for action: some behavioral and neurophysiological considerations of attention and action. In: Heuer H, Sanders A.F, editors. Perspectives on perception and action. Lawrence Erlbaum Associates; Hillsdale, NJ: 1987. pp. 395–419.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources