

Human immunodeficiency virus type 1 variants resistant to first- and second-version fusion inhibitors and cytopathic in ex vivo human lymphoid tissue
- PMID: 17428857
- PMCID: PMC1900115
- DOI: 10.1128/JVI.02546-06
Human immunodeficiency virus type 1 variants resistant to first- and second-version fusion inhibitors and cytopathic in ex vivo human lymphoid tissue
Abstract
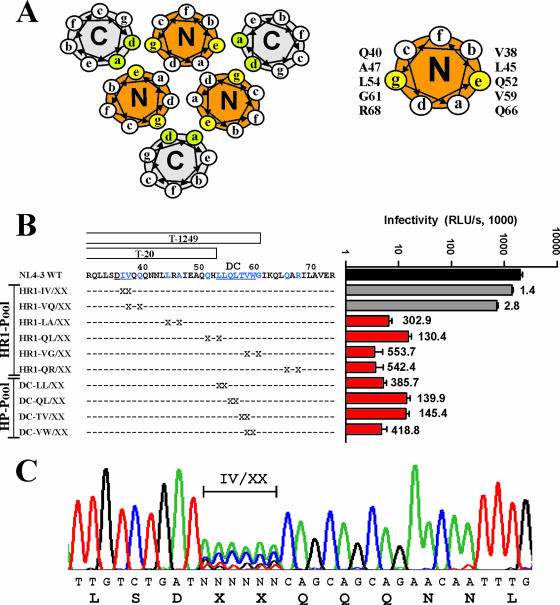
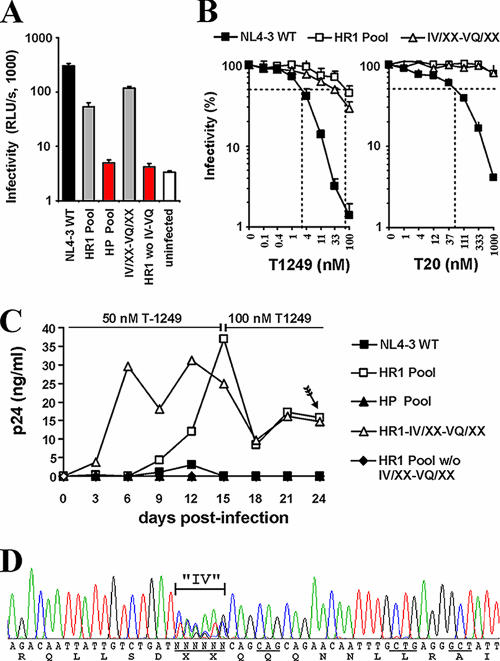
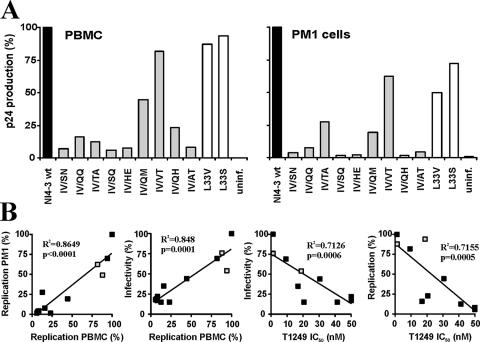
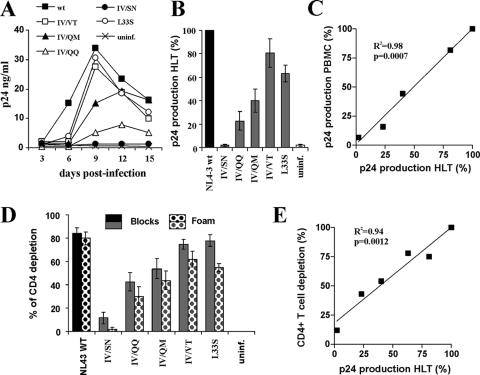
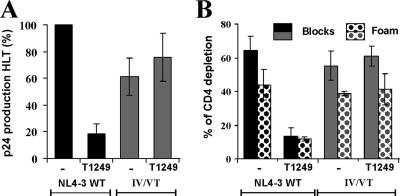
Human immunodeficiency virus type 1 (HIV-1) fusion inhibitors blocking viral entry by binding the gp41 heptad repeat 1 (HR1) region offer great promise for antiretroviral therapy, and the first of these inhibitors, T20 (Fuzeon; enfuvirtide), is successfully used in the clinic. It has been reported previously that changes in the 3-amino-acid GIV motif at positions 36 to 38 of gp41 HR1 mediate resistance to T20 but usually not to second-version fusion inhibitors, such as T1249, which target an overlapping but distinct region in HR1 including a conserved hydrophobic pocket (HP). Based on the common lack of cross-resistance and the difficulty of selecting T1249-resistant HIV-1 variants, it has been suggested that the determinants of resistance to first- and second-version fusion inhibitors may be different. To further assess HIV-1 resistance to fusion inhibitors and to analyze where changes in HR1 are tolerated, we randomized 16 codons in the HR1 region, including those making contact with HR2 codons and/or encoding residues in the GIV motif and the HP. We found that changes only at positions 37I, 38V, and 40Q near the N terminus of HR1 were tolerated. The propagation of randomly gp41-mutated HIV-1 variants in the presence of T1249 allowed the effective selection of highly resistant forms, all containing changes in the IV residues. Overall, the extent of T1249 resistance was inversely correlated to viral fitness and cytopathicity. Notably, one HIV-1 mutant showing approximately 10-fold-reduced susceptibility to T1249 inhibition replicated with wild type-like kinetics and caused substantial CD4+-T-cell depletion in ex vivo-infected human lymphoid tissue in the presence and absence of an inhibitor. Taken together, our results show that the GIV motif also plays a key role in resistance to second-version fusion inhibitors and suggest that some resistant HIV-1 variants may be pathogenic in vivo.
Figures
Similar articles
-
Heptad-repeat-2 mutations enhance the stability of the enfuvirtide-resistant HIV-1 gp41 hairpin structure.Antivir Ther. 2005;10(8):893-900. Antivir Ther. 2005. PMID: 16430194
-
Selection of T1249-resistant human immunodeficiency virus type 1 variants.J Virol. 2008 Jul;82(13):6678-88. doi: 10.1128/JVI.00352-08. Epub 2008 Apr 23. J Virol. 2008. PMID: 18434391 Free PMC article.
-
HIV-1 gp41 Residues Modulate CD4-Induced Conformational Changes in the Envelope Glycoprotein and Evolution of a Relaxed Conformation of gp120.J Virol. 2018 Jul 31;92(16):e00583-18. doi: 10.1128/JVI.00583-18. Print 2018 Aug 15. J Virol. 2018. PMID: 29875245 Free PMC article.
-
HIV-1 gp41 heptad repeat 2 (HR2) possesses an amino acid domain that resembles the allergen domain in Aspergillus fumigatus Asp f1 protein: review, hypothesis and implications.Virus Genes. 2007 Jun;34(3):233-40. doi: 10.1007/s11262-007-0082-3. Epub 2007 Mar 1. Virus Genes. 2007. PMID: 17333401 Review.
-
Synthesized peptide inhibitors of HIV-1 gp41-dependent membrane fusion.Curr Pharm Des. 2013;19(10):1800-9. doi: 10.2174/1381612811319100004. Curr Pharm Des. 2013. PMID: 23092277 Review.
Cited by
-
Combinations of the first and next generations of human immunodeficiency virus (HIV) fusion inhibitors exhibit a highly potent synergistic effect against enfuvirtide- sensitive and -resistant HIV type 1 strains.J Virol. 2009 Aug;83(16):7862-72. doi: 10.1128/JVI.00168-09. Epub 2009 Jun 3. J Virol. 2009. PMID: 19493996 Free PMC article.
-
Structural basis of potent and broad HIV-1 fusion inhibitor CP32M.J Biol Chem. 2012 Aug 3;287(32):26618-29. doi: 10.1074/jbc.M112.381079. Epub 2012 Jun 7. J Biol Chem. 2012. PMID: 22679024 Free PMC article.
-
Resistance of human immunodeficiency virus type 1 to a third-generation fusion inhibitor requires multiple mutations in gp41 and is accompanied by a dramatic loss of gp41 function.J Virol. 2011 Oct;85(20):10785-97. doi: 10.1128/JVI.05331-11. Epub 2011 Aug 10. J Virol. 2011. PMID: 21835789 Free PMC article.
-
Optimization of peptidic HIV-1 fusion inhibitor T20 by phage display.Protein Sci. 2019 Aug;28(8):1501-1512. doi: 10.1002/pro.3669. Protein Sci. 2019. PMID: 31228294 Free PMC article.
-
Biophysical property and broad anti-HIV activity of albuvirtide, a 3-maleimimidopropionic acid-modified peptide fusion inhibitor.PLoS One. 2012;7(3):e32599. doi: 10.1371/journal.pone.0032599. Epub 2012 Mar 5. PLoS One. 2012. PMID: 22403678 Free PMC article.
References
-
- Agrawal, L., X. Lu, Q. Jin, and G. G. Alkhatib. 2006. Anti-HIV therapy: current and future directions. Curr. Pharm. Des. 12:2031-2055. - PubMed
-
- Aquaro, S., R. D'Arrigo, V. Svicher, G. Perri, S. L. Caputo, U. Visco-Comandini, M. Santoro, A. Bertoli, F. Mazzotta, S. Bonora, V. Tozzi, R. Bellagamba, M. Zaccarelli, P. Narciso, and C. F. Antinori. 2006. Specific mutations in HIV-1 gp41 are associated with immunological success in HIV-1-infected patients receiving enfuvirtide treatment. J. Antimicrob. Chemother. 58:714-722. - PubMed
-
- Armand-Ugon, M., A. Gutierrez, B. Clotet, and E. A. Este. 2003. HIV-1 resistance to the gp41-dependent fusion inhibitor C-34. Antivir. Res. 59:137-142. - PubMed
-
- Briz, V., E. Poveda, and V. Soriano. 2006. HIV entry inhibitors: mechanisms of action and resistance pathways. J. Antimicrob. Chemother. 57:619-627. - PubMed
-
- Carl, S., R. Daniels, A. J. Iafrate, P. Easterbrook, T. C. Greenough, J. Skowronski, and F. Kirchhoff. 2000. Partial “repair” of defective nef genes in a long-term nonprogressor with human immunodeficiency virus type 1 infection. J. Infect. Dis. 181:132-140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
