

Persistent binding of ligands to the aryl hydrocarbon receptor
- PMID: 17431010
- PMCID: PMC2857992
- DOI: 10.1093/toxsci/kfm085
Persistent binding of ligands to the aryl hydrocarbon receptor
Abstract
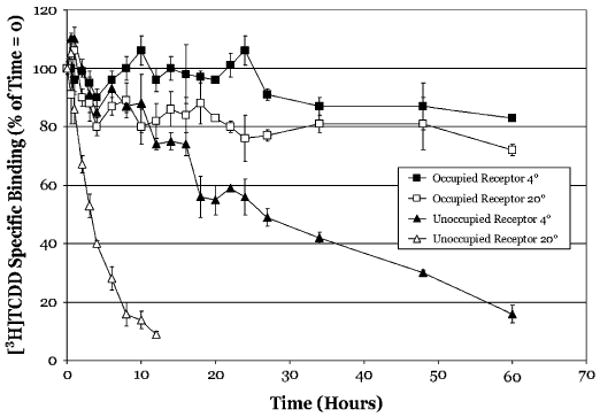
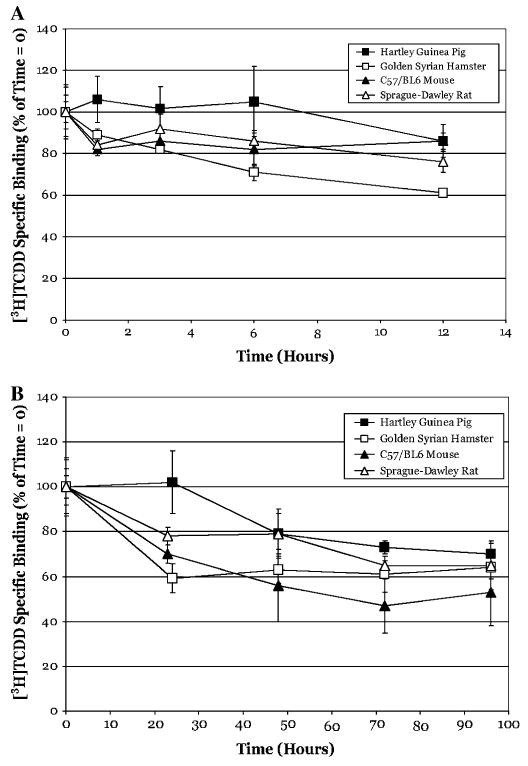
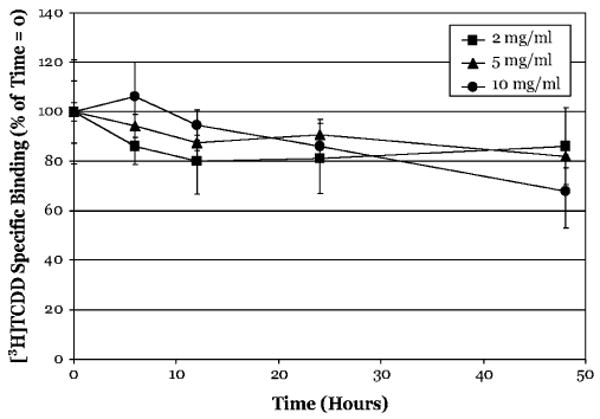
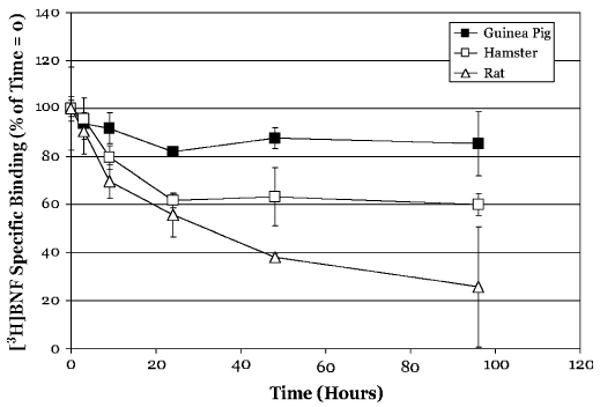
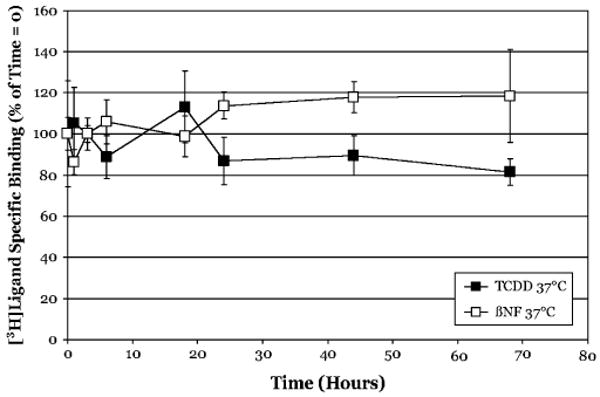
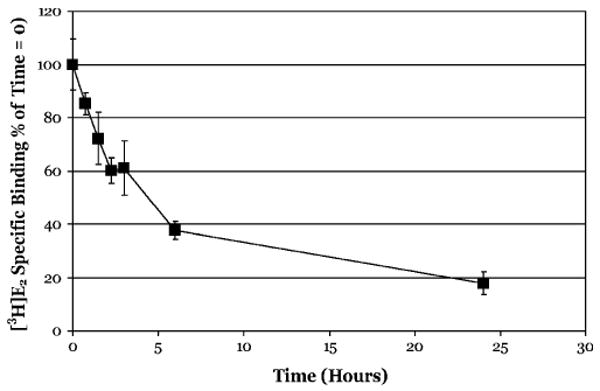
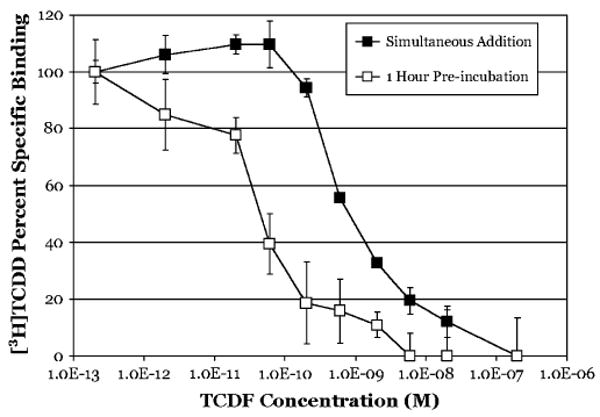
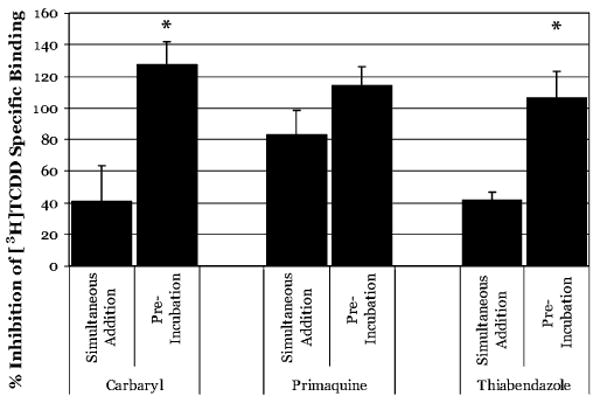
The aryl hydrocarbon receptor (AhR) is a ligand-activated transcription factor that mediates many of the biological and toxic effects of halogenated aromatic hydrocarbons (HAHs), polycyclic aromatic hydrocarbons (PAHs), and other structurally diverse ligands. While HAHs are several orders of magnitude more potent in producing AhR-dependent biochemical effects than PAHs or other AhR agonists, only the HAHs have been observed to produce AhR-dependent toxicity in vivo. Here we have characterized the dissociation of a prototypical HAH ligand ([(3)H] 2,3,7,8-tetrachlorodibenzo-p-dioxin [TCDD]) and PAH-like ligand ([(3)H] beta-naphthoflavone [betaNF]) from the guinea pig, hamster, mouse, and rat hepatic cytosolic AhR in order to elucidate the relationship between the apparent ligand-binding affinities and the divergent potency of these chemicals. Both compounds dissociated very slowly from the AhR with the amount of specific binding remaining at 96 h ranging from 53% to 70% for [(3)H]TCDD and 26% to 85% for [(3)H] betaNF, depending upon the species examined. The rate of ligand dissociation was unaffected by protein concentration or incubation temperature. Preincubation of cytosol with 2,3,7,8-tetrachlorodibenzofuran, carbaryl, or primaquine, prior to the addition of [(3)H]TCDD, shifted the apparent IC(50) of these compounds as competitive AhR ligands by approximately 10- to 50-fold. Our results support the need for reassessment of previous AhR ligand-binding affinity calculations and competitive binding analysis since these measurements are not carried out at equilibrium binding conditions. Our studies suggest that AhR binding affinity/occupancy has little effect on the observed differences in the persistence of gene expression by HAHs and PAHs.
Figures
References
-
- Backlund M, Ingelman-Sundberg M. Different structural requirements of the ligand binding domain of the aryl hydrocarbon receptor for high- and low-affinity ligand binding and receptor activation. Mol Pharmacol. 2004;65:416–425. - PubMed
-
- Bank PA, Yao EF, Phelps CL, Harper PA, Denison MS. Species-specific binding of transformed Ah receptor to a dioxin responsive transcriptional enhancer. Eur J Pharmacol. 1992;228:85–94. - PubMed
-
- Berghard A, Gradin K, Toftgard R. The stability of dioxin-receptor ligands influences cytochrome P450IA1 expression in human keratinocytes. Carcinogenesis. 1992;13:651–655. - PubMed
-
- Boctor AM, Band P, Grossman A. Analysis of binding of [3H]Estradiol to the cytosol fraction of rat pancreas: Comparison with sites in the cytosol of uterus. Endocrinology. 1983;113:453–462. - PubMed
-
- Bradfield CA, Kende AS, Poland A. Kinetic and equilibrium studies of Ah receptor-ligand binding: Use of [125I]2-iodo-7,8-dibromodibenzo-p-dioxin. Mol Pharmacol. 1988;34:229–237. - PubMed
