

XTsh3 is an essential enhancing factor of canonical Wnt signaling in Xenopus axial determination
- PMID: 17431396
- PMCID: PMC1864982
- DOI: 10.1038/sj.emboj.7601684
XTsh3 is an essential enhancing factor of canonical Wnt signaling in Xenopus axial determination
Abstract
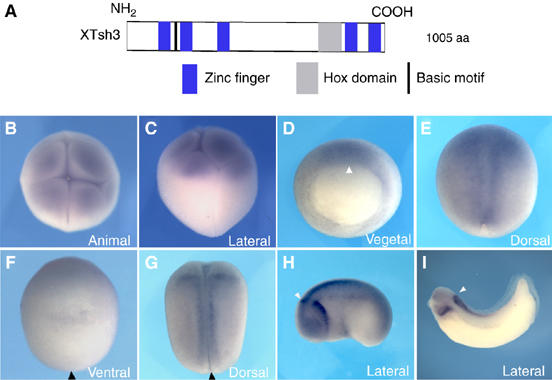
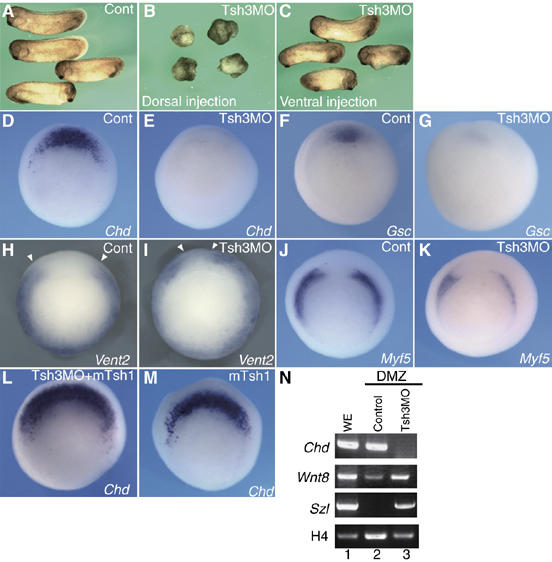
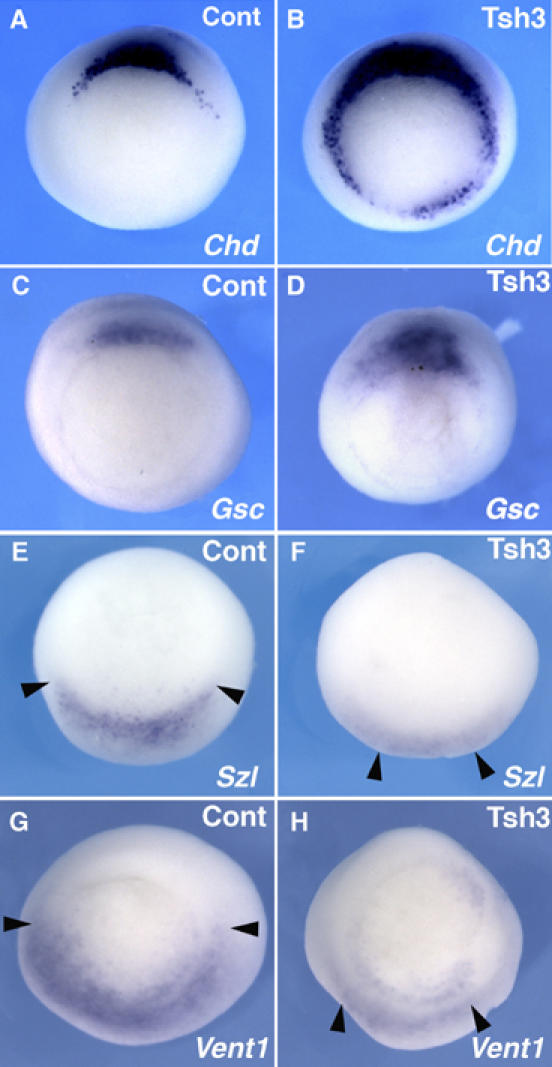
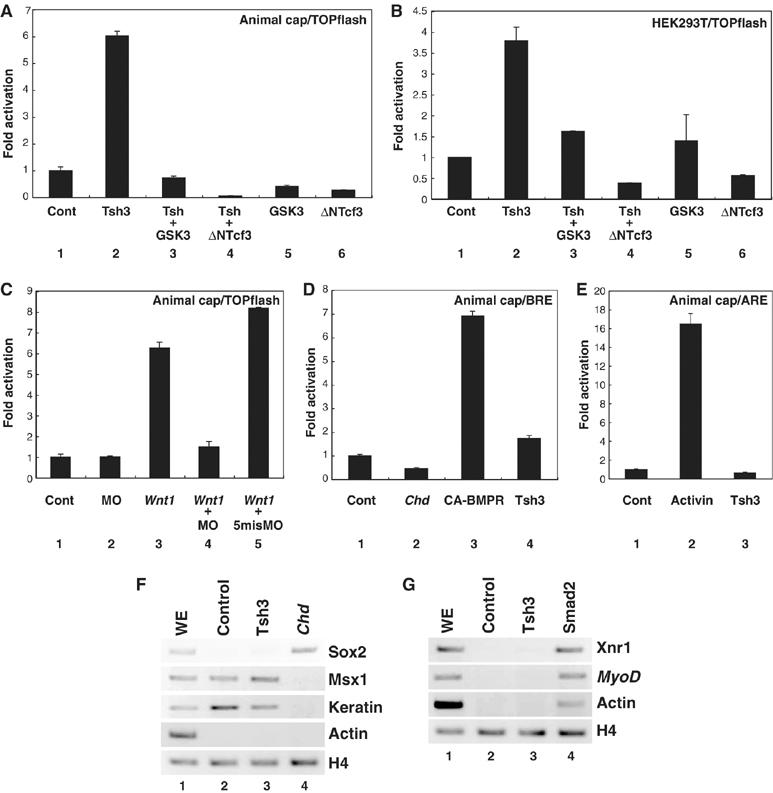
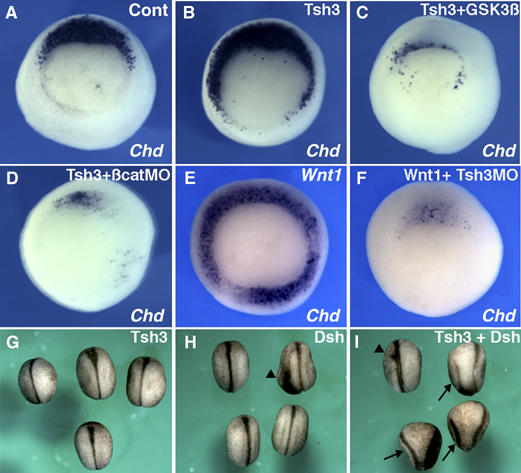
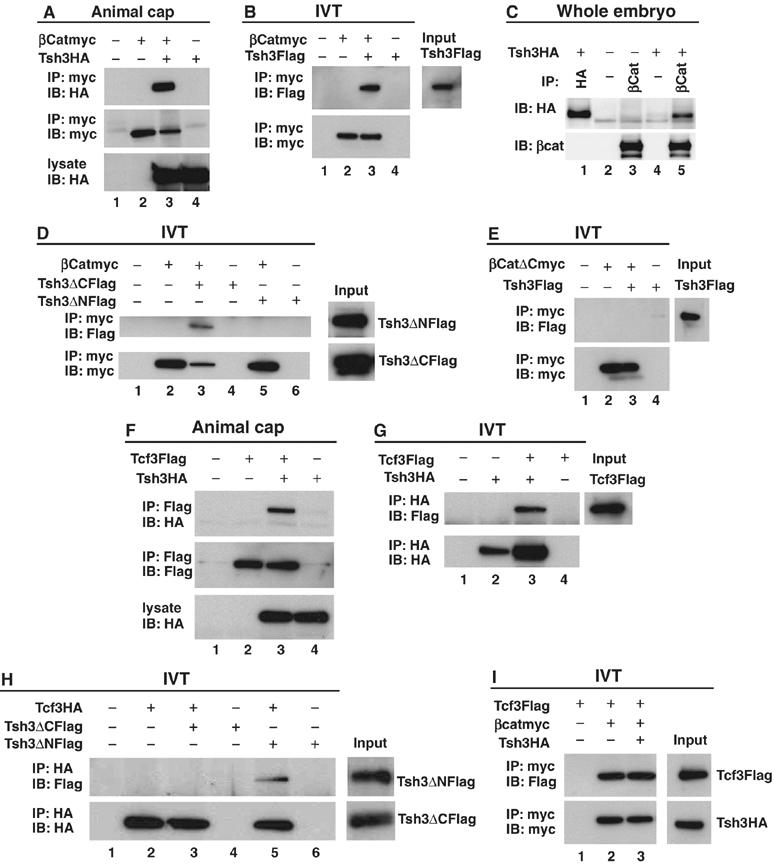
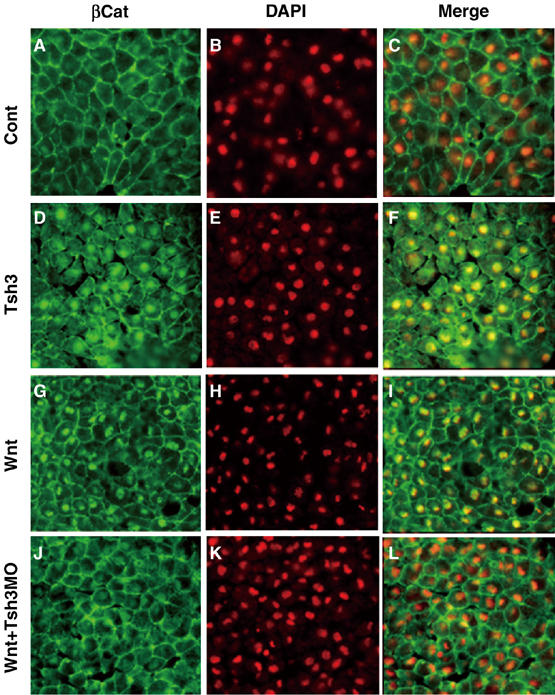
In Xenopus, an asymmetric distribution of Wnt activity that follows cortical rotation in the fertilized egg leads to the dorsal-ventral (DV) axis establishment. However, how a clear DV polarity develops from the initial difference in Wnt activity still remains elusive. We report here that the Teashirt-class Zn-finger factor XTsh3 plays an essential role in dorsal determination by enhancing canonical Wnt signaling. Knockdown of the XTsh3 function causes ventralization in the Xenopus embryo. Both in vivo and in vitro studies show that XTsh3 substantially enhances Wnt signaling activity in a beta-catenin-dependent manner. XTsh3 cooperatively promotes the formation of a secondary axis on the ventral side when combined with weak Wnt activity, whereas XTsh3 alone has little axis-inducing ability. Furthermore, Wnt1 requires XTsh3 for its dorsalizing activity in vivo. Immunostaining and protein analyses indicate that XTsh3 is a nuclear protein that physically associates with beta-catenin and efficiently increases the level of beta-catenin in the nucleus. We discuss the role of XTsh3 as an essential amplifying factor of canonical Wnt signaling in embryonic dorsal determination.
Figures
Similar articles
-
Neural induction in Xenopus requires inhibition of Wnt-beta-catenin signaling.Dev Biol. 2006 Oct 1;298(1):71-86. doi: 10.1016/j.ydbio.2006.06.015. Epub 2006 Jun 14. Dev Biol. 2006. PMID: 16879817
-
Traf2- and Nck-interacting kinase is essential for canonical Wnt signaling in Xenopus axis formation.J Biol Chem. 2010 Aug 20;285(34):26289-94. doi: 10.1074/jbc.M109.090597. Epub 2010 Jun 21. J Biol Chem. 2010. PMID: 20566648 Free PMC article.
-
Jun NH2-terminal kinase (JNK) prevents nuclear beta-catenin accumulation and regulates axis formation in Xenopus embryos.Proc Natl Acad Sci U S A. 2006 Oct 31;103(44):16313-8. doi: 10.1073/pnas.0602557103. Epub 2006 Oct 23. Proc Natl Acad Sci U S A. 2006. PMID: 17060633 Free PMC article.
-
The canonical Wnt pathway in embryonic axis polarity.Semin Cell Dev Biol. 2006 Apr;17(2):168-74. doi: 10.1016/j.semcdb.2006.04.004. Epub 2006 Apr 25. Semin Cell Dev Biol. 2006. PMID: 16714126 Review.
-
Emergence of organizer function: a lot of "stuff" involved.Semin Cell Dev Biol. 2006 Feb;17(1):110-6. doi: 10.1016/j.semcdb.2006.01.002. Epub 2006 Feb 28. Semin Cell Dev Biol. 2006. PMID: 16503175 Review.
Cited by
-
Teashirt-3, a novel regulator of muscle differentiation, associates with BRG1-associated factor 57 (BAF57) to inhibit myogenin gene expression.J Biol Chem. 2011 Jul 1;286(26):23498-510. doi: 10.1074/jbc.M110.206003. Epub 2011 May 4. J Biol Chem. 2011. PMID: 21543328 Free PMC article.
-
Ancestral mesodermal reorganization and evolution of the vertebrate head.Zoological Lett. 2015 Nov 9;1:29. doi: 10.1186/s40851-015-0030-3. eCollection 2015. Zoological Lett. 2015. PMID: 26605074 Free PMC article.
-
Go ahead, grow a head! A planarian's guide to anterior regeneration.Regeneration (Oxf). 2016 Jun 24;3(3):139-55. doi: 10.1002/reg2.56. eCollection 2016 Jun. Regeneration (Oxf). 2016. PMID: 27606065 Free PMC article. Review.
-
Bmp4 Synexpression Gene, Sizzled, Transcription Is Collectively Modulated by Smad1 and Ventx1.1/Ventx2.1 in Early Xenopus Embryos.Int J Mol Sci. 2022 Nov 1;23(21):13335. doi: 10.3390/ijms232113335. Int J Mol Sci. 2022. PMID: 36362118 Free PMC article.
-
teashirt is required for head-versus-tail regeneration polarity in planarians.Development. 2015 Mar 15;142(6):1062-72. doi: 10.1242/dev.119685. Epub 2015 Feb 27. Development. 2015. PMID: 25725068 Free PMC article.
References
-
- Abe K, Takeichi M (2007) NMDA-receptor activation induces calpain-mediated beta-catenin cleavages for triggering gene expression. Neuron 53: 387–397 - PubMed
-
- Bessa J, Casares F (2005) Restricted teashirt expression confers eye-specific responsiveness to Dpp and Wg signals during eye specification in Drosophila. Development 132: 5011–5020 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
