

Interleukin-4 impairs granzyme-mediated cytotoxicity of Simian virus 40 large tumor antigen-specific CTL in BALB/c mice
- PMID: 17431618
- PMCID: PMC11030854
- DOI: 10.1007/s00262-007-0309-0
Interleukin-4 impairs granzyme-mediated cytotoxicity of Simian virus 40 large tumor antigen-specific CTL in BALB/c mice
Abstract
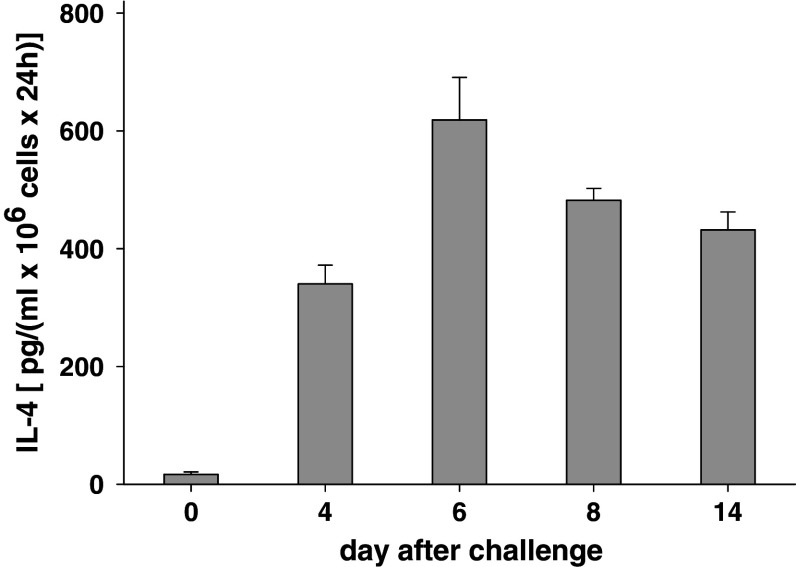
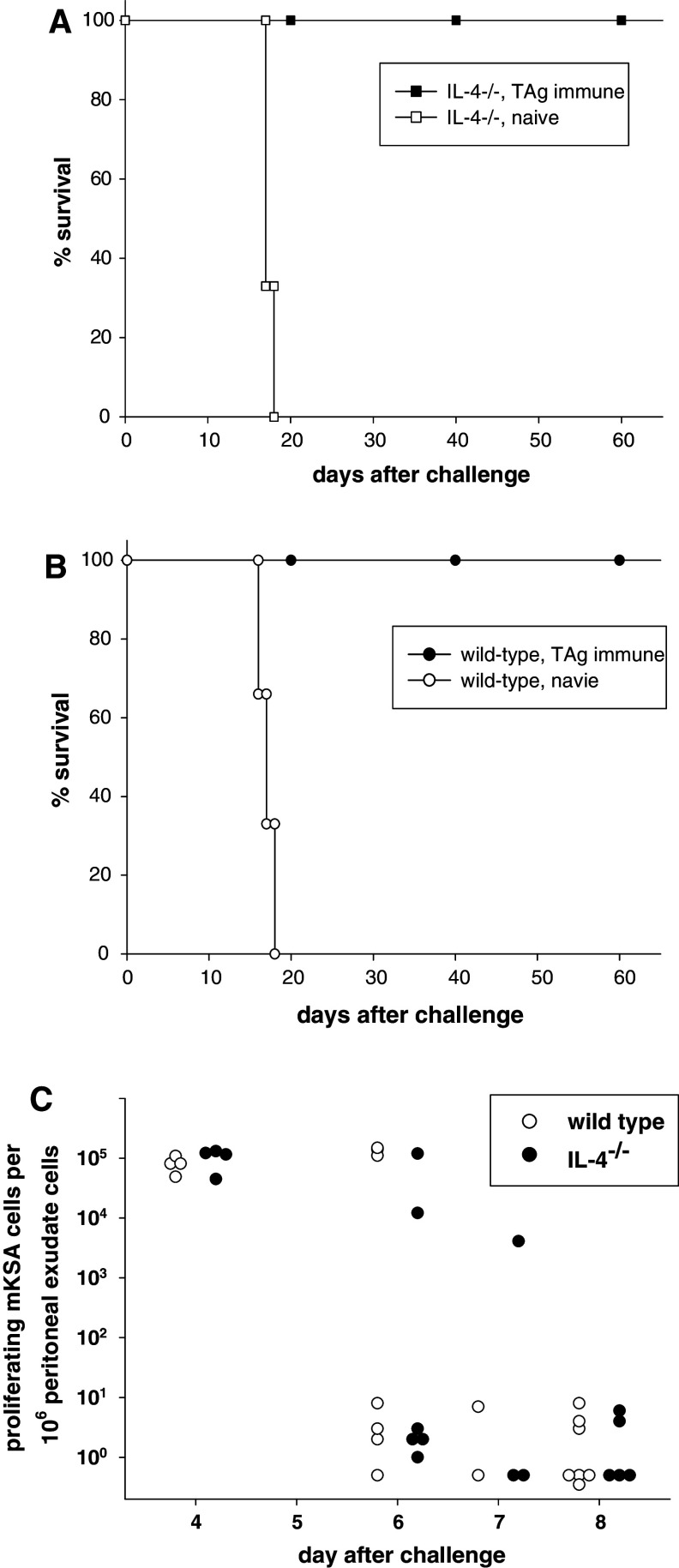
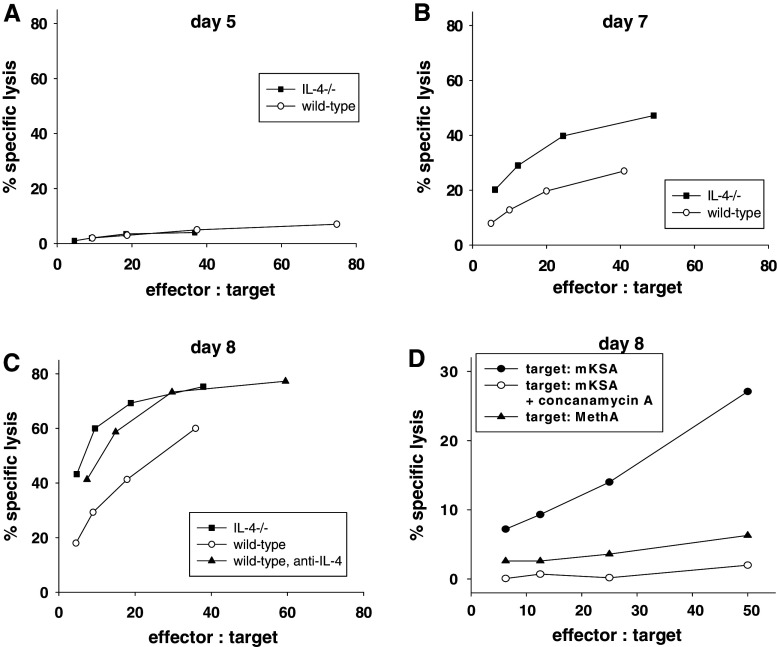
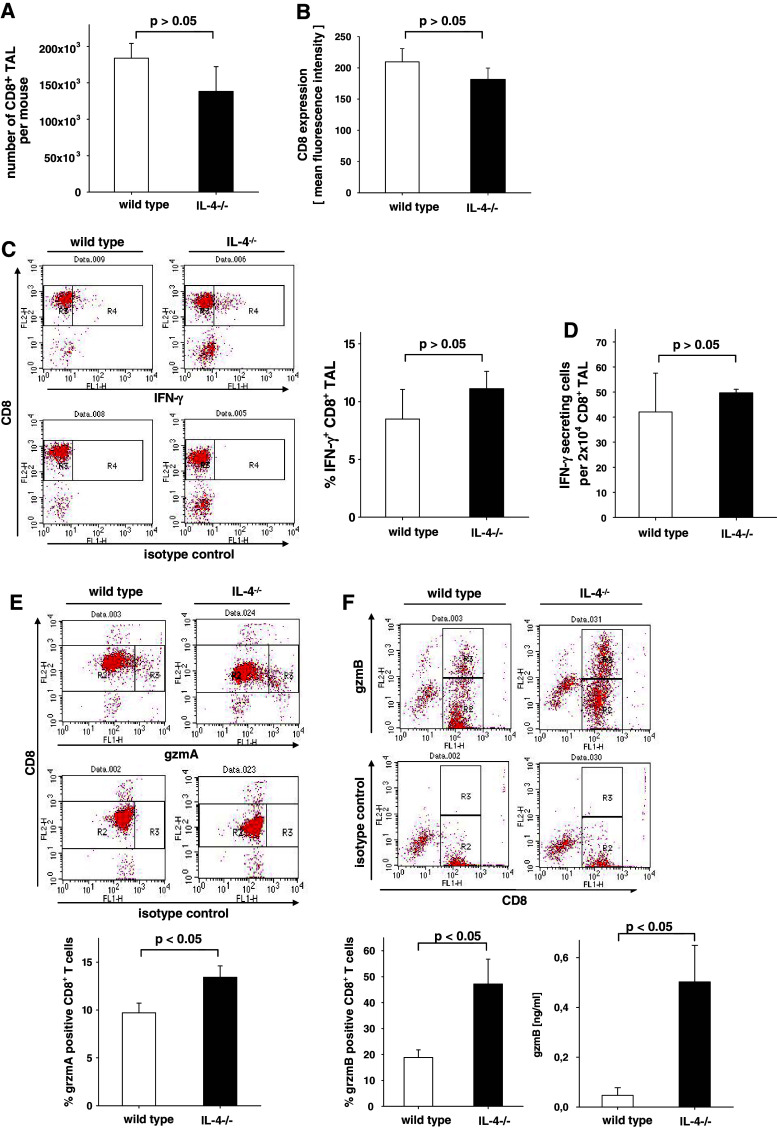
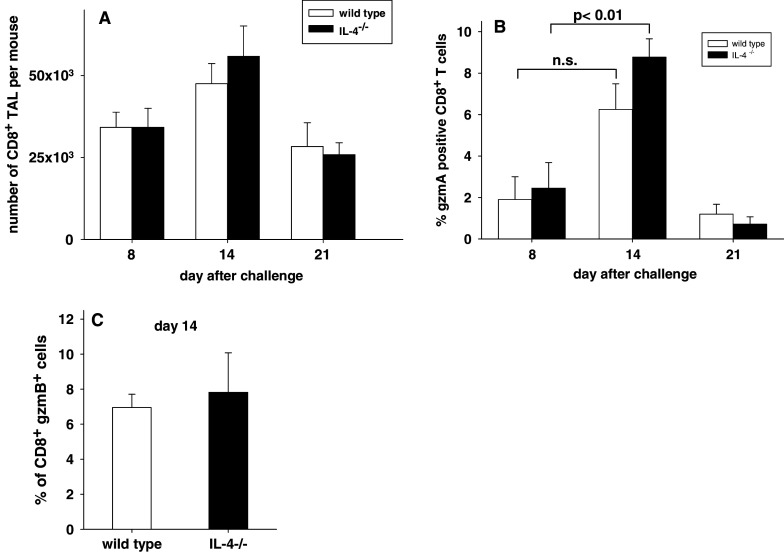
In this report we analyzed the impact of interleukin-4 (IL-4) on tumor-associated simian virus 40 (SV40) large T-antigen (TAg)-specific CD8+ cytotoxic T cells during rejection of syngeneic SV40 transformed mKSA tumor cells in BALB/c mice. Strikingly, challenge of naïve mice with low doses of mKSA tumor cells revealed a CD8+ T cell-dependent prolonged survival time of naïve IL-4-/- mice. In mice immunized with SV40 TAg we observed in IL-4-/- mice, or in wild type mice treated with neutralizing anti-IL-4 monoclonal antibody, a strongly enhanced TAg-specific cytotoxicity of tumor associated CD8+ T cells. The enhanced cytotoxicity in IL-4-/- mice was accompanied by a significant increase in the fraction of CD8+ tumor associated T-cells expressing the cytotoxic effector molecules granzyme A and B and in granzyme B-specific enzymatic activity. The data suggest that endogenous IL-4 can suppress the generation of CD8+ CTL expressing cytotoxic effector molecules especially when the antigen induces only a very weak CTL response.
Figures

Similar articles
-
Tumor immunity against a simian virus 40 oncoprotein requires CD8+ T lymphocytes in the effector immune phase.J Virol. 2010 Jan;84(2):883-93. doi: 10.1128/JVI.01512-09. Epub 2009 Nov 4. J Virol. 2010. PMID: 19889780 Free PMC article.
-
Simian virus 40 large-T-antigen-specific rejection of mKSA tumor cells in BALB/c mice is critically dependent on both strictly tumor-associated, tumor-specific CD8(+) cytotoxic T lymphocytes and CD4(+) T helper cells.J Virol. 2001 Nov;75(22):10593-602. doi: 10.1128/JVI.75.22.10593-10602.2001. J Virol. 2001. PMID: 11602701 Free PMC article.
-
Role of a subdominant H-2Kd-restricted SV40 tumor antigen cytotoxic T lymphocyte epitope in tumor rejection.Virology. 1998 May 10;244(2):427-41. doi: 10.1006/viro.1998.9148. Virology. 1998. PMID: 9601511
-
Immunization of BALB/c mice with recombinant simian virus 40 large tumor antigen induces antibody-dependent cell-mediated cytotoxicity against simian virus 40-transformed cells. An antibody-based mechanism for tumor immunity.J Immunol. 1994 Sep 1;153(5):2064-71. J Immunol. 1994. PMID: 8051411
-
Recognition of simian virus 40 T antigen by cytotoxic T lymphocytes.Mol Biol Med. 1990 Feb;7(1):83-96. Mol Biol Med. 1990. PMID: 2157938 Review.
Cited by
-
Role of the innate immune response and tumor immunity associated with simian virus 40 large tumor antigen.J Virol. 2010 Oct;84(19):10121-30. doi: 10.1128/JVI.01095-10. Epub 2010 Jul 28. J Virol. 2010. PMID: 20668083 Free PMC article.
-
Tumor immunity against a simian virus 40 oncoprotein requires CD8+ T lymphocytes in the effector immune phase.J Virol. 2010 Jan;84(2):883-93. doi: 10.1128/JVI.01512-09. Epub 2009 Nov 4. J Virol. 2010. PMID: 19889780 Free PMC article.
-
Alternative activation of tumor-associated macrophages by IL-4: priming for protumoral functions.Cell Cycle. 2010 Dec 15;9(24):4824-35. doi: 10.4161/cc.9.24.14322. Epub 2010 Dec 15. Cell Cycle. 2010. PMID: 21150330 Free PMC article. Review.
-
Interleukin-4-induced loss of CD8 expression and cytolytic function in effector CD8 T cells persists long term in vivo.Immunology. 2013 Jun;139(2):187-96. doi: 10.1111/imm.12068. Immunology. 2013. PMID: 23311920 Free PMC article.
-
Interleukin-4 suppresses the cytotoxic potential of in vitro generated, adaptive regulatory CD4 T cells by down-regulation of granzyme B.Immunology. 2009 Jul;127(3):338-44. doi: 10.1111/j.1365-2567.2008.02993.x. Epub 2008 Dec 23. Immunology. 2009. PMID: 19191911 Free PMC article.
References
-
- Trenn G, Takayama H, Hu-Li J, Paul WE, Sitkovsky MV. B cell stimulatory factor 1 (IL-4) enhances the development of cytotoxic T cells from Lyt-2+ resting murine T lymphocytes. J Immunol. 1988;140:1101–1106. - PubMed
-
- Miller CL, Hooton JW, Gillis S, Paetkau V. IL-4 potentiates the IL-2-dependent proliferation of mouse cytotoxic T cells. J Immunol. 1990;144:1331–1337. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
