

Signalling within the neurovascular unit in the mammalian retina
- PMID: 17434916
- PMCID: PMC2279186
- DOI: 10.1113/expphysiol.2006.036376
Signalling within the neurovascular unit in the mammalian retina
Abstract
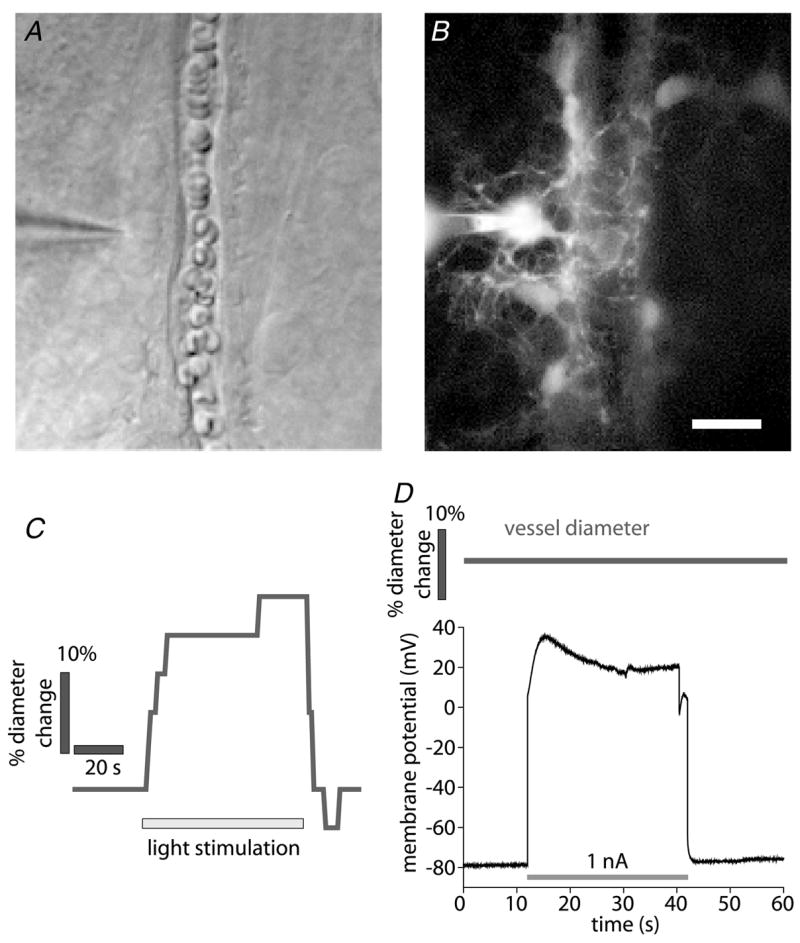
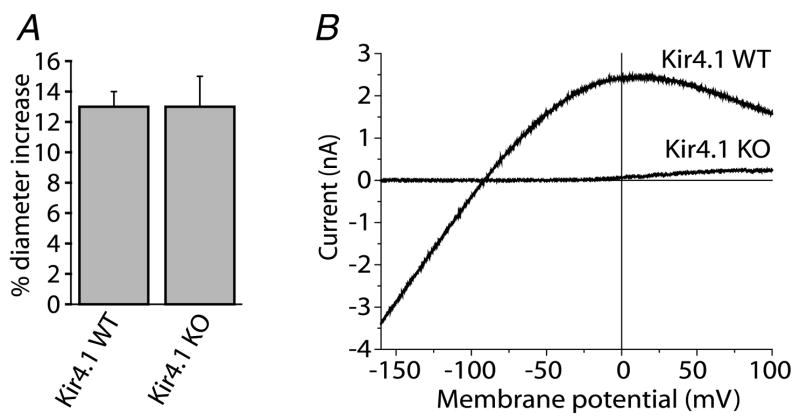
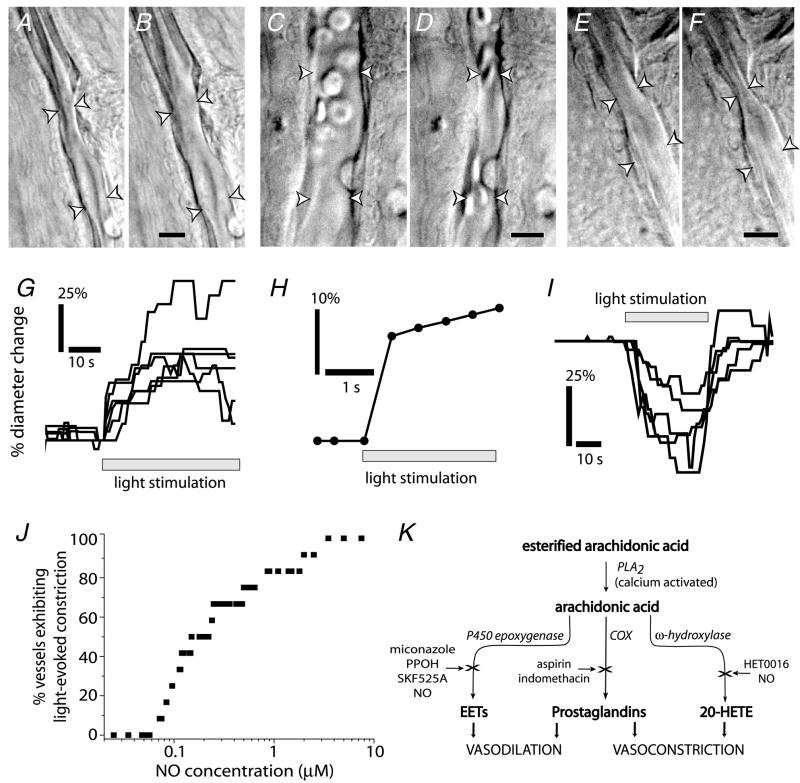
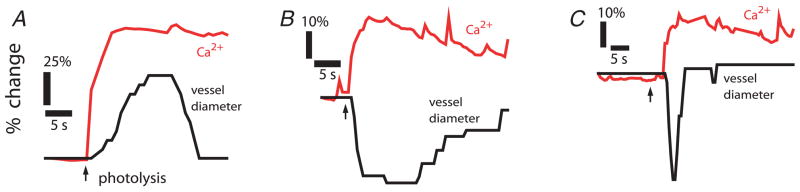
Neuronal activity in the central nervous system evokes localized changes in blood flow, a response termed neurovascular coupling or functional hyperaemia. Modern functional imaging methods, such as functional magnetic resonance imaging (fMRI), measure signals related to functional hyperaemia in order to determine localization of brain function and to diagnose disease. The cellular mechanisms that underlie functional hyperaemia, however, are not well understood. Glial cells have been hypothesized to be intermediaries between neurons and blood vessels in the control of neurovascular coupling, owing to their ability to release vasoactive factors in response to neuronal activity. Using an in vitro preparation of the isolated, intact rodent retina, we have investigated two likely mechanisms of glial control of the vasculature: glial K(+) siphoning and glial induction of vasoactive arachidonic acid metabolites. Potassium siphoning is a process by which a K(+) current flowing through glial cells transfers K(+) released from active neurons to blood vessels. Since slight increases in extracellular K(+) can cause vasodilatation, this mechanism was hypothesized to contribute to neurovascular coupling. Our data, however, suggest that glial K(+) siphoning does not contribute significantly to neurovascular coupling in the retina. Instead, we suggest that glial cells mediate neurovascular coupling by inducing the production of two types of arachidonic acid metabolites, epoxyeicosatrienoic acids (EETs) and 20-hydroxyeicosatetraenoic acid (20-HETE), which dilate and constrict vessels, respectively. We show that both light flashes and direct glial stimulation produce vasodilatation or vasoconstriction mediated by EETs and 20-HETE, respectively. Further, we show that the type of vasomotor response observed (dilatation or constriction) depends on retinal levels of nitric oxide. Our data also demonstrate that glial cells are necessary intermediaries for signalling from neurons to blood vessels, since functional hyperaemia does not occur when neuron-to-glia communication is interrupted. These results indicate that glial cells play an important role in mediating functional hyperaemia and suggest that the regulation of blood flow may involve both vasodilating and vasoconstricting components.
Figures
References
-
- Badeer HS. Hemodynamics for medical students. Adv Physiol Educ. 2001;25:44–52. - PubMed
-
- Brew H, Gray PTA, Mobbs P, Attwell D. Endfeet of retinal glial cells have higher densities of ion channels that mediate K+ buffering. Nature. 1986;324:466–468. - PubMed
-
- Chrissobolis S, Sobey CG. Inwardly rectifying potassium channels in the regulation of vascular tone. Curr Drug Targets. 2003;4:281–289. - PubMed
-
- Chrissobolis S, Ziogas J, Chu Y, Faraci FM, Sobey CG. Role of inwardly rectifying K+ channels in K+-induced cerebral vasodilatation in vivo. Am J Physiol Heart Circ Physiol. 2000;279:H2704–H2712. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources