

Inhibition of cyclooxygenases by dipyrone
- PMID: 17435797
- PMCID: PMC2013970
- DOI: 10.1038/sj.bjp.0707239
Inhibition of cyclooxygenases by dipyrone
Abstract
Background and purpose: Dipyrone is a potent analgesic drug that has been demonstrated to inhibit cyclooxygenase (COX). In contrast to classical COX-inhibitors, such as aspirin-like drugs, dipyrone has no anti-inflammatory effect and a low gastrointestinal toxicity, indicating a different mode of action. Here, we aimed to investigate the effects of dipyrone on COX.
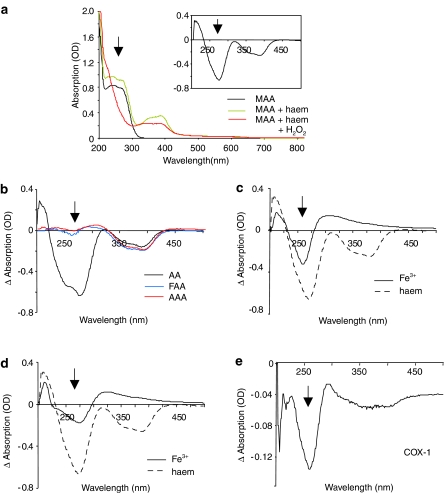
Experimental approach: The four major metabolites of dipyrone, including the two pharmacologically active metabolites, 4-methyl-amino-antipyrine (MAA) and amino-antipyrine (AA), were used to characterise their binding to COX and haem as well as their effects on the biochemical properties of COX. Mass spectrometry, UV and visible photometry were used to study binding and prostaglandin production. Levels of anti-oxidant enzymes were assessed by Western blotting.
Key results: The pharmacologically active metabolites of dipyrone, MAA and AA, did not inhibit COX activity in vitro like classical COX inhibitors, but instead redirected the prostaglandin synthesis, ruling out inhibition of COX through binding to its active site. We found that MAA and AA formed stable complexes with haem and reacted with hydrogen peroxide in presence of haem, ferrous ions (Fe(2+)) or COX. Moreover, MAA reduced Fe(3+) to Fe(2+) and accordingly increased lipid peroxidation and the expression of anti-oxidant enzymes in cultured cells and in vivo.
Conclusions and implications: Our data suggest that the pharmacologically active metabolites of dipyrone inhibit COX activity by sequestering radicals which initiate the catalytic activity of this enzyme or through the reduction of the oxidative states of the COX protein.
Figures




References
-
- Abbate R, Gori AM, Pinto S, Attanasio M, Paniccia R, Coppo M, et al. Cyclooxygenase and lipoxygenase metabolite synthesis by polymorphonuclear neutrophils: in vitro effect of dipyrone. Prostaglandins Leukot Essent Fatty Acids. 1990;41:89–93. - PubMed
-
- Aronoff DM, Boutaud O, Marnett LJ, Oates JA. Inhibition of prostaglandin H2 synthases by salicylate is dependent on the oxidative state of the enzymes. J Pharmacol Exp Ther. 2003;304:589–595. - PubMed
-
- Brenneis C, Maier TJ, Schmidt R, Hofacker A, Zulauf L, Jakobsson PJ, et al. Inhibition of prostaglandin E2 synthesis by SC-560 is independent of cyclooxygenase 1 inhibition. FASEB J. 2006;20:1352–1360. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
