

A subset of dendritic cells induces CD4+ T cells to produce IFN-gamma by an IL-12-independent but CD70-dependent mechanism in vivo
- PMID: 17438065
- PMCID: PMC2118574
- DOI: 10.1084/jem.20070176
A subset of dendritic cells induces CD4+ T cells to produce IFN-gamma by an IL-12-independent but CD70-dependent mechanism in vivo
Abstract
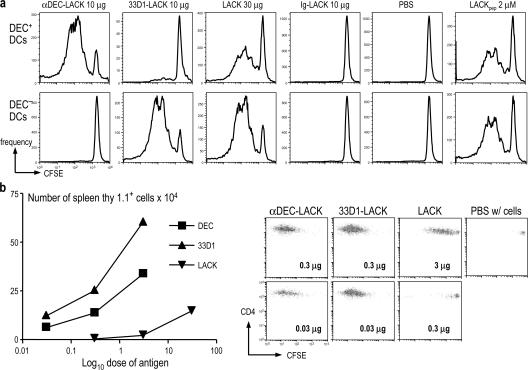
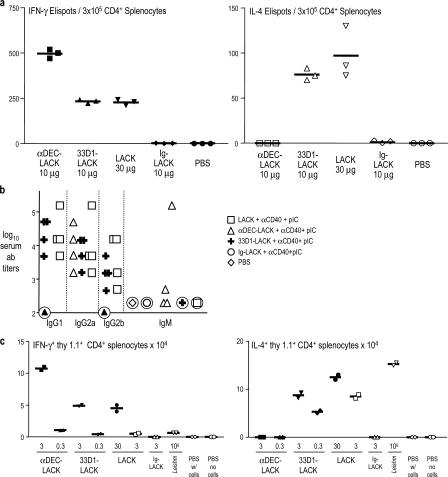
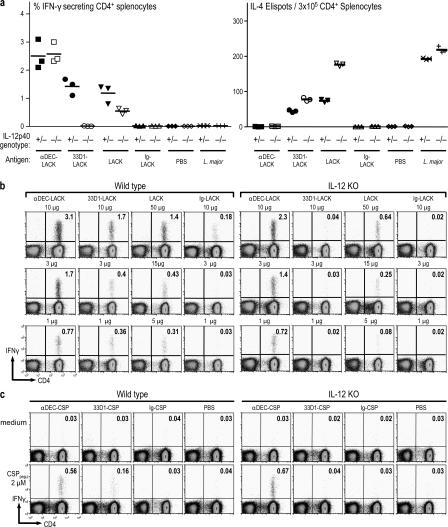
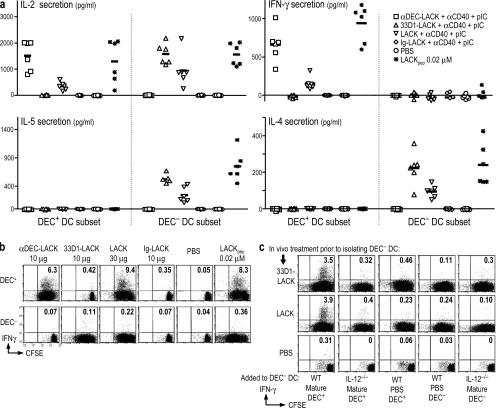
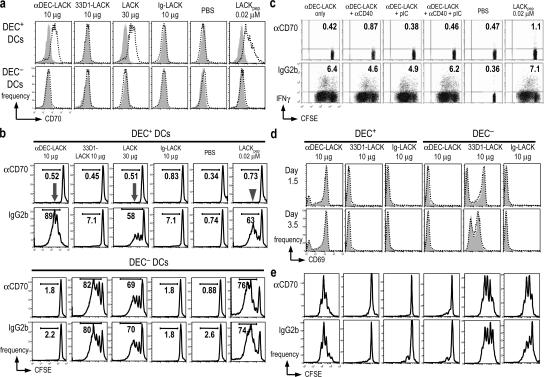
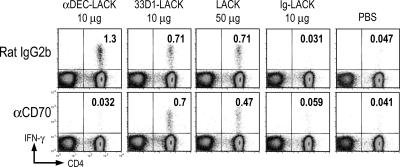
Interferon (IFN)-gamma, a cytokine critical for resistance to infection and tumors, is produced by CD4(+) helper T lymphocytes after stimulation by cultured dendritic cells (DCs) that secrete a cofactor, interleukin (IL)-12. We have identified a major IL-12-independent pathway whereby DCs induce IFN-gamma-secreting T helper (Th)1 CD4(+) T cells in vivo. This pathway requires the membrane-associated tumor necrosis family member CD70 and was identified by targeting the LACK antigen from Leishmania major within an antibody to CD205 (DEC-205), an uptake receptor on a subset of DCs. Another major DC subset, targeted with 33D1 anti-DCIR2 antibody, also induced IFN-gamma in vivo but required IL-12, not CD70. Isolated CD205(+) DCs expressed cell surface CD70 when presenting antigen to T cell receptor transgenic T cells, and this distinction was independent of maturation stimuli. CD70 was also essential for CD205(+) DC function in vivo. Detection of the IL-12-independent IFN-gamma pathway was obscured with nontargeted LACK, which was presented by both DC subsets. This in situ analysis points to CD70 as a decision maker for Th1 differentiation by CD205(+) DCs, even in Th2-prone BALB/c animals and potentially in vaccine design. The results indicate that two DC subsets have innate propensities to differentially affect the Th1/Th2 balance in vivo and by distinct mechanisms.
Figures
References
-
- Abbas, A.K., K.M. Murphy, and A. Sher. 1996. Functional diversity of helper T lymphocytes. Nature. 383:787–793. - PubMed
-
- O'Garra, A. 1998. Cytokines induce the development of functionally heterogeneous T helper cell subsets. Immunity. 8:275–283. - PubMed
-
- Murphy, K.M., and S.L. Reiner. 2002. The lineage decisions of helper T cells. Nat. Rev. Immunol. 2:933–944. - PubMed
-
- Shankaran, V., H. Ikeda, A.T. Bruce, J.M. White, P.E. Swanson, L.J. Old, and R.D. Schreiber. 2001. IFNγ and lymphocytes prevent primary tumour development and shape tumour immunogenicity. Nature. 410:1107–1111. - PubMed
-
- Mosmann, T.R., and R.L. Coffman. 1989. Th1 and Th2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 7:145–173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
