

Microphthalmia transcription factor isoforms in mast cells and the heart
- PMID: 17438132
- PMCID: PMC1900032
- DOI: 10.1128/MCB.01455-06
Microphthalmia transcription factor isoforms in mast cells and the heart
Abstract
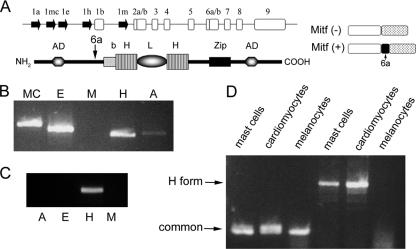
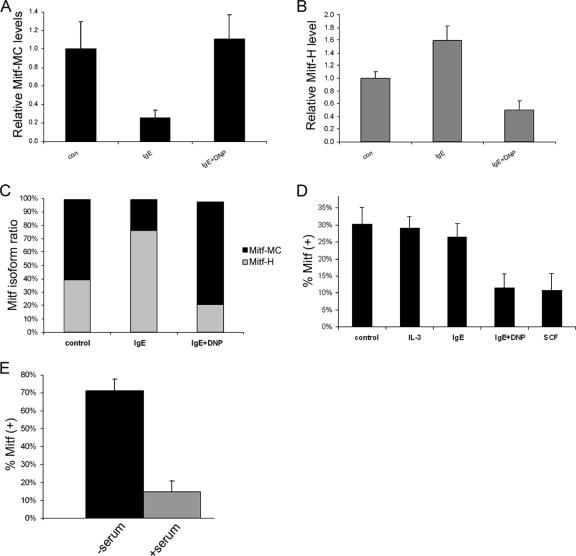
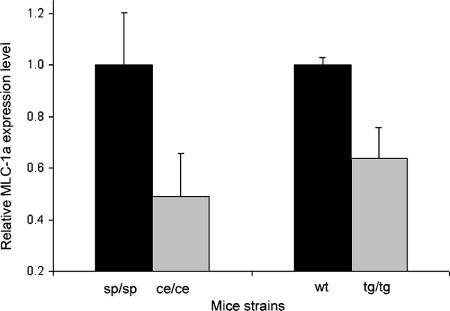
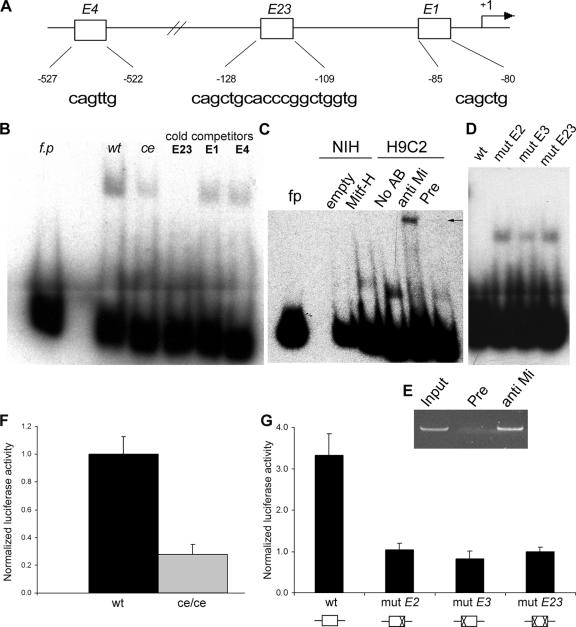
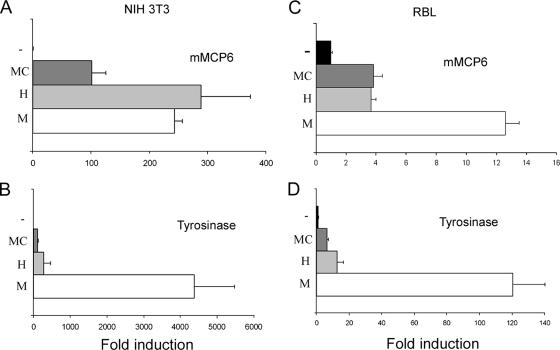
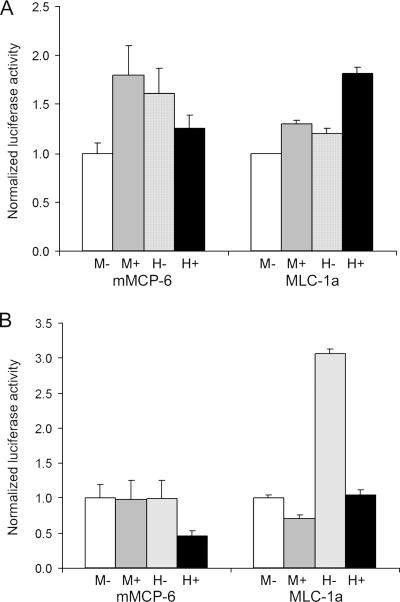
The microphthalmia transcription factor (Mitf) is critical for the survival and differentiation of a variety of cell types. While on the transcript level it has been noted that melanocytes and cardiomyocytes express specific Mitf isoforms, mast cells express several isoforms, mainly Mitf-H and Mitf-MC, whose function has not been thoroughly investigated. We found that in mast cells the expression of the specific Mitf isoforms is dependent on physiological stimuli that cause a major shifting of promoter usage and internal splicing. For example, activation of the c-kit signaling pathway almost totally abolished one of the main splice isoforms. Since cardiomyocytes express only the Mitf-H isoform, they were an ideal system to determine this isoform's physiological role. We identified that the expression of myosin light-chain 1a (MLC-1a) is regulated by Mitf-H. Interestingly, the transactivation of MLC-1a by Mitf-H in cardiomyocytes is decreased by overexpression of the splice form with exon 6a. In conclusion, we found that there is physiological switching of Mitf isoforms and that the promoter context and the cell context have a combined influence on gene expression programs.
Figures

References
-
- Bischoff, S. C., and G. Sellge. 2002. Mast cell hyperplasia: role of cytokines. Int. Arch. Allergy Immunol. 127:118-122. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases