

High incidence of non-random template strand segregation and asymmetric fate determination in dividing stem cells and their progeny
- PMID: 17439301
- PMCID: PMC1852584
- DOI: 10.1371/journal.pbio.0050102
High incidence of non-random template strand segregation and asymmetric fate determination in dividing stem cells and their progeny
Erratum in
- PLoS Biol. 2007 Jul;5(7):e182
Abstract
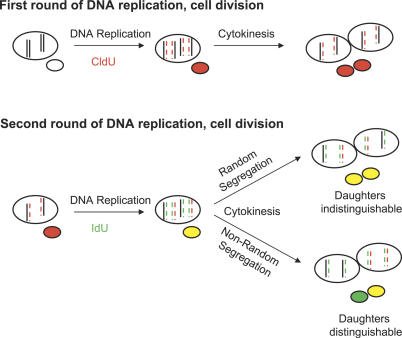
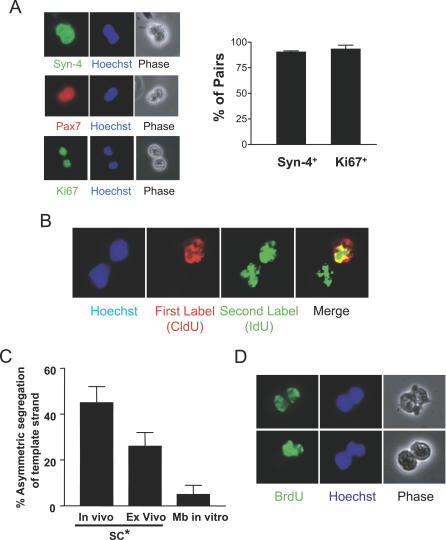
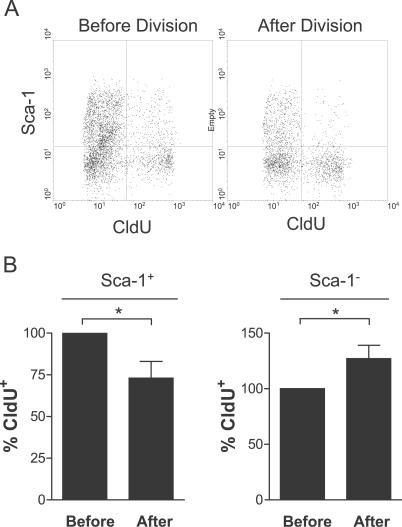
Decades ago, the "immortal strand hypothesis" was proposed as a means by which stem cells might limit acquiring mutations that could give rise to cancer, while continuing to proliferate for the life of an organism. Originally based on observations in embryonic cells, and later studied in terms of stem cell self-renewal, this hypothesis has remained largely unaccepted because of few additional reports, the rarity of the cells displaying template strand segregation, and alternative interpretations of experiments involving single labels or different types of labels to follow template strands. Using sequential pulses of halogenated thymidine analogs (bromodeoxyuridine [BrdU], chlorodeoxyuridine [CldU], and iododeoxyuridine [IdU]), and analyzing stem cell progeny during induced regeneration in vivo, we observed extraordinarily high frequencies of segregation of older and younger template strands during a period of proliferative expansion of muscle stem cells. Furthermore, template strand co-segregation was strongly associated with asymmetric cell divisions yielding daughters with divergent fates. Daughter cells inheriting the older templates retained the more immature phenotype, whereas daughters inheriting the newer templates acquired a more differentiated phenotype. These data provide compelling evidence of template strand co-segregation based on template age and associated with cell fate determination, suggest that template strand age is monitored during stem cell lineage progression, and raise important caveats for the interpretation of label-retaining cells.
Conflict of interest statement
Figures


Comment in
-
Non-random chromosome segregation in stem cells.PLoS Biol. 2007 May;5(5):e125. doi: 10.1371/journal.pbio.0050125. Epub 2007 Apr 17. PLoS Biol. 2007. PMID: 20076670 Free PMC article. No abstract available.
References
-
- Cairns J. Mutation selection and the natural history of cancer. Nature. 1975;255:197–200. - PubMed
-
- Lark KG, Consigli RA, Minocha HC. Segregation of sister chromatids in mammalian cells. Science. 1966;154:1202–1205. - PubMed
-
- Potten CS, Hume WJ, Reid P, Cairns J. The segregation of DNA in epithelial stem cells. Cell. 1978;15:899–906. - PubMed
-
- Merok JR, Lansita JA, Tunstead JR, Sherley JL. Cosegregation of chromosomes containing immortal DNA strands in cells that cycle with asymmetric stem cell kinetics. Cancer Res. 2002;62:6791–6795. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
