

Peptides encoded by short ORFs control development and define a new eukaryotic gene family
- PMID: 17439302
- PMCID: PMC1852585
- DOI: 10.1371/journal.pbio.0050106
Peptides encoded by short ORFs control development and define a new eukaryotic gene family
Abstract
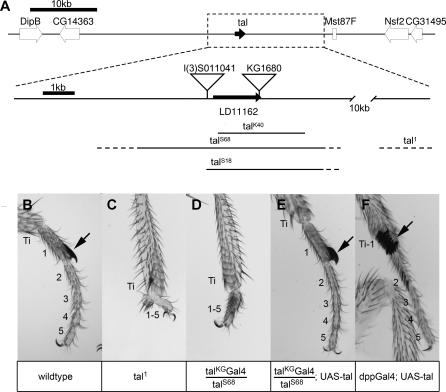
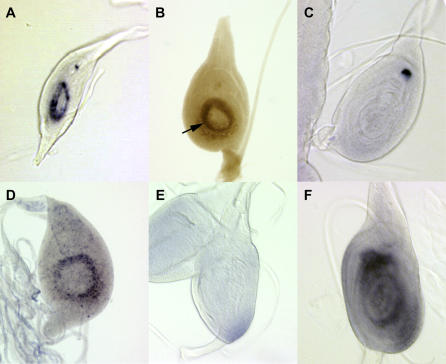
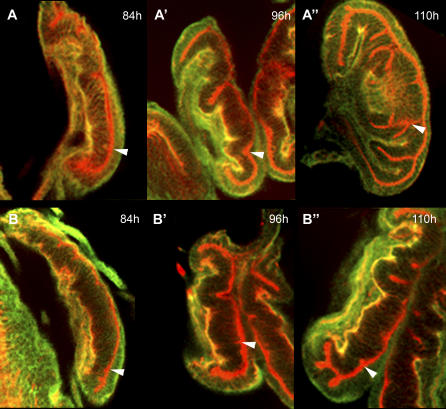
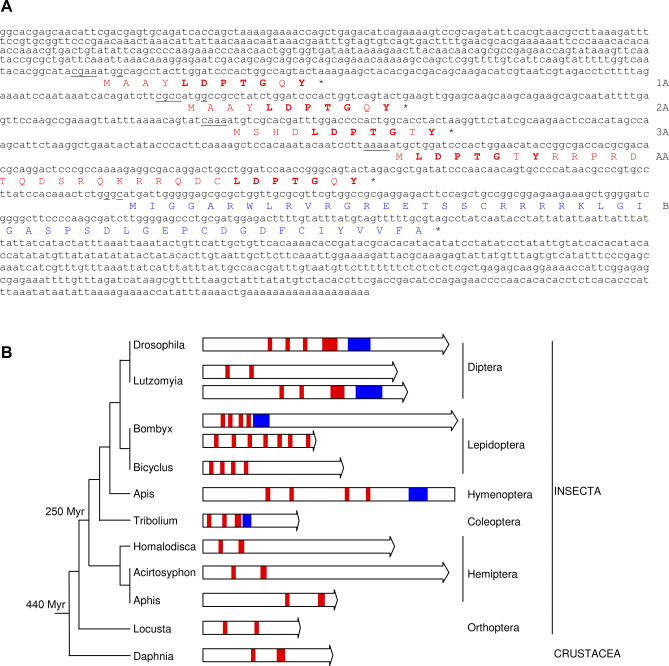
Despite recent advances in developmental biology, and the sequencing and annotation of genomes, key questions regarding the organisation of cells into embryos remain. One possibility is that uncharacterised genes having nonstandard coding arrangements and functions could provide some of the answers. Here we present the characterisation of tarsal-less (tal), a new type of noncanonical gene that had been previously classified as a putative noncoding RNA. We show that tal controls gene expression and tissue folding in Drosophila, thus acting as a link between patterning and morphogenesis. tal function is mediated by several 33-nucleotide-long open reading frames (ORFs), which are translated into 11-amino-acid-long peptides. These are the shortest functional ORFs described to date, and therefore tal defines two novel paradigms in eukaryotic coding genes: the existence of short, unprocessed peptides with key biological functions, and their arrangement in polycistronic messengers. Our discovery of tal-related short ORFs in other species defines an ancient and noncanonical gene family in metazoans that represents a new class of eukaryotic genes. Our results open a new avenue for the annotation and functional analysis of genes and sequenced genomes, in which thousands of short ORFs are still uncharacterised.
Conflict of interest statement
Figures


References
-
- Consortium IHGS. Finishing the euchromatic sequence of the human genome. Nature. 2004;431:931–945. - PubMed
-
- Pollard KS, Salama SR, Lambert N, Lambot M-A, Coppens S, et al. An RNA gene expressed during cortical development evolved rapidly in humans. Nature. 2006;443:167–172. - PubMed
-
- Couso JP, Bishop SA. Proximo-distal development in the legs of Drosophila . Int J Dev Biol. 1998;42:345–352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
