

On the functions of the h subunit of eukaryotic initiation factor 3 in late stages of translation initiation
- PMID: 17439654
- PMCID: PMC1896003
- DOI: 10.1186/gb-2007-8-4-r60
On the functions of the h subunit of eukaryotic initiation factor 3 in late stages of translation initiation
Abstract
Background: The eukaryotic translation initiation factor 3 (eIF3) has multiple roles during the initiation of translation of cytoplasmic mRNAs. How individual subunits of eIF3 contribute to the translation of specific mRNAs remains poorly understood, however. This is true in particular for those subunits that are not conserved in budding yeast, such as eIF3h.
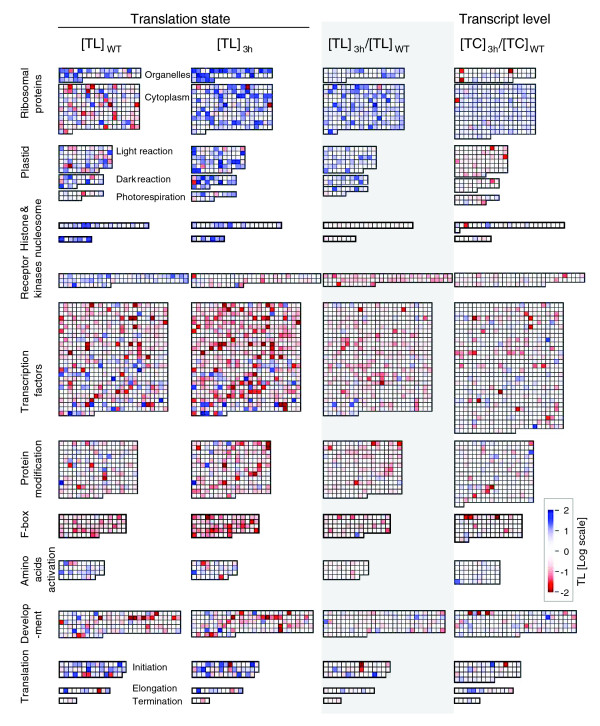
Results: Working with stable reporter transgenes in Arabidopsis thaliana mutants, it was demonstrated that the h subunit of eIF3 contributes to the efficient translation initiation of mRNAs harboring upstream open reading frames (uORFs) in their 5' leader sequence. uORFs, which can function as devices for translational regulation, are present in over 30% of Arabidopsis mRNAs, and are enriched among mRNAs for transcriptional regulators and protein modifying enzymes. Microarray comparisons of polysome loading in wild-type and eif3h mutant seedlings revealed that eIF3h generally helps to maintain efficient polysome loading of mRNAs harboring multiple uORFs. In addition, however, eIF3h also boosted the polysome loading of mRNAs with long leaders or coding sequences. Moreover, the relative polysome loading of certain functional groups of mRNAs, including ribosomal proteins, was actually increased in the eif3h mutant, suggesting that regulons of translational control can be revealed by mutations in generic translation initiation factors.
Conclusion: The intact eIF3h protein contributes to efficient translation initiation on 5' leader sequences harboring multiple uORFs, although mRNA features independent of uORFs are also implicated.
Figures









Similar articles
-
Translational regulation via 5' mRNA leader sequences revealed by mutational analysis of the Arabidopsis translation initiation factor subunit eIF3h.Plant Cell. 2004 Dec;16(12):3341-56. doi: 10.1105/tpc.104.026880. Epub 2004 Nov 17. Plant Cell. 2004. PMID: 15548739 Free PMC article.
-
The h subunit of eIF3 promotes reinitiation competence during translation of mRNAs harboring upstream open reading frames.RNA. 2010 Apr;16(4):748-61. doi: 10.1261/rna.2056010. Epub 2010 Feb 23. RNA. 2010. PMID: 20179149 Free PMC article.
-
Translation reinitiation and development are compromised in similar ways by mutations in translation initiation factor eIF3h and the ribosomal protein RPL24.BMC Plant Biol. 2010 Aug 27;10:193. doi: 10.1186/1471-2229-10-193. BMC Plant Biol. 2010. PMID: 20799971 Free PMC article.
-
Does eIF3 promote reinitiation after translation of short upstream ORFs also in mammalian cells?RNA Biol. 2017 Dec 2;14(12):1660-1667. doi: 10.1080/15476286.2017.1353863. Epub 2017 Sep 15. RNA Biol. 2017. PMID: 28745933 Free PMC article. Review.
-
Regulation of plant translation by upstream open reading frames.Plant Sci. 2014 Jan;214:1-12. doi: 10.1016/j.plantsci.2013.09.006. Epub 2013 Sep 18. Plant Sci. 2014. PMID: 24268158 Review.
Cited by
-
RNA around the clock - regulation at the RNA level in biological timing.Front Plant Sci. 2015 May 5;6:311. doi: 10.3389/fpls.2015.00311. eCollection 2015. Front Plant Sci. 2015. PMID: 25999975 Free PMC article. Review.
-
Interaction of PABPC1 with the translation initiation complex is critical to the NMD resistance of AUG-proximal nonsense mutations.Nucleic Acids Res. 2012 Feb;40(3):1160-73. doi: 10.1093/nar/gkr820. Epub 2011 Oct 11. Nucleic Acids Res. 2012. PMID: 21989405 Free PMC article.
-
TOR tour to auxin.EMBO J. 2013 Apr 17;32(8):1069-71. doi: 10.1038/emboj.2013.69. Epub 2013 Mar 22. EMBO J. 2013. PMID: 23524852 Free PMC article.
-
A Transcript-Specific eIF3 Complex Mediates Global Translational Control of Energy Metabolism.Cell Rep. 2016 Aug 16;16(7):1891-902. doi: 10.1016/j.celrep.2016.07.006. Epub 2016 Jul 28. Cell Rep. 2016. PMID: 27477275 Free PMC article.
-
Translation initiation factor eIF3h targets specific transcripts to polysomes during embryogenesis.Proc Natl Acad Sci U S A. 2013 Jun 11;110(24):9818-23. doi: 10.1073/pnas.1302934110. Epub 2013 May 28. Proc Natl Acad Sci U S A. 2013. PMID: 23716667 Free PMC article.
References
-
- Unbehaun A, Borukhov SI, Hellen CU, Pestova TV. Release of initiation factors from 48S complexes during ribosomal subunit joining and the link between establishment of codon-anticodon base-pairing and hydrolysis of eIF2-bound GTP. Genes Dev. 2004;18:3078–3093. doi: 10.1101/gad.1255704. - DOI - PMC - PubMed
-
- Dong Z, Zhang JT. Initiation factor eIF3 and regulation of mRNA translation, cell growth, and cancer. Crit Rev Oncol Hematol. 2006;59:169–180. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
