

p53 and p21 determine the sensitivity of noscapine-induced apoptosis in colon cancer cells
- PMID: 17440101
- PMCID: PMC3757339
- DOI: 10.1158/0008-5472.CAN-06-4282
p53 and p21 determine the sensitivity of noscapine-induced apoptosis in colon cancer cells
Abstract
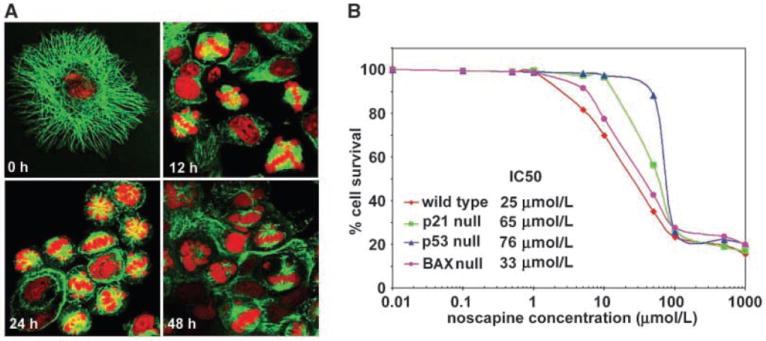
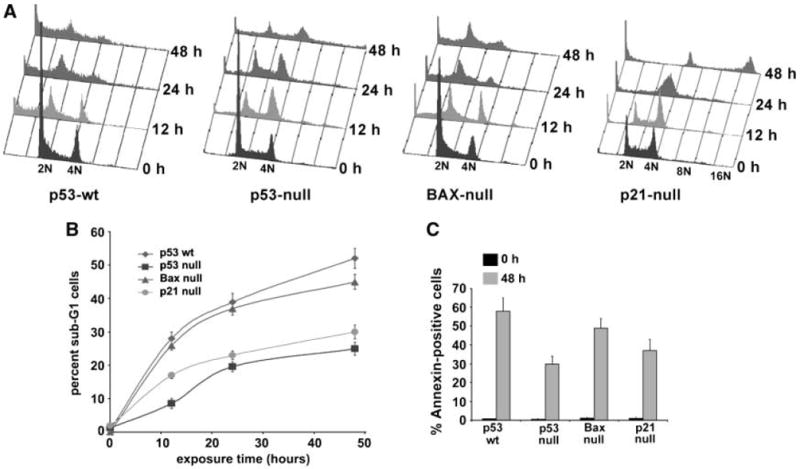
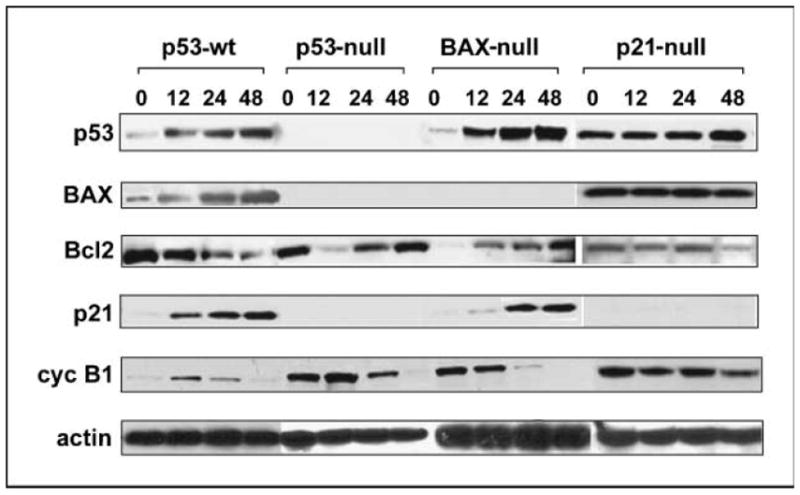
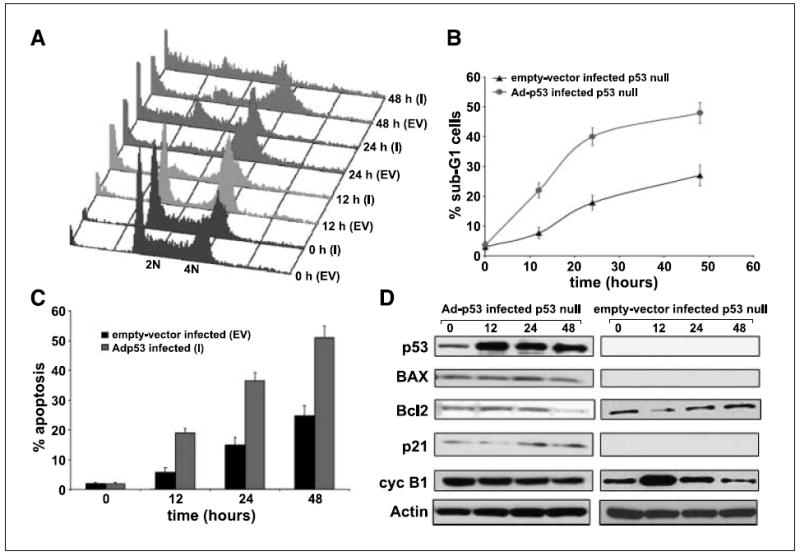
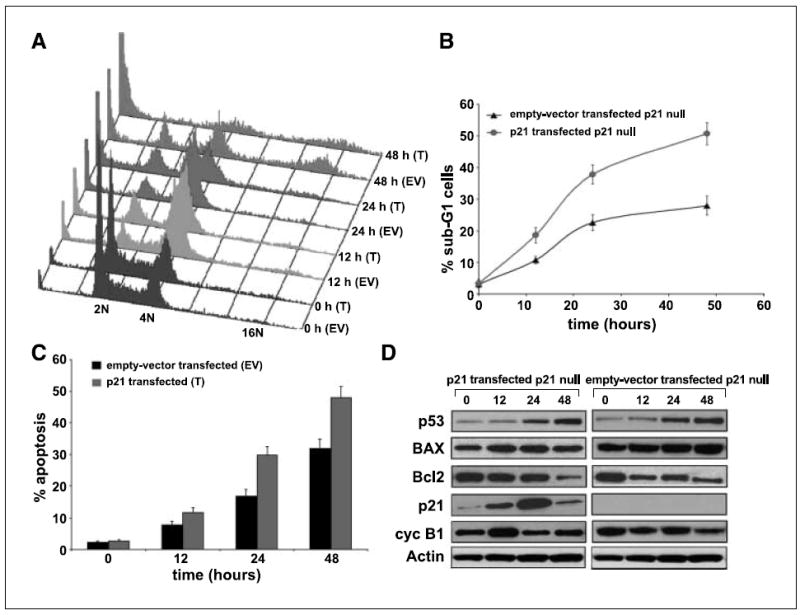
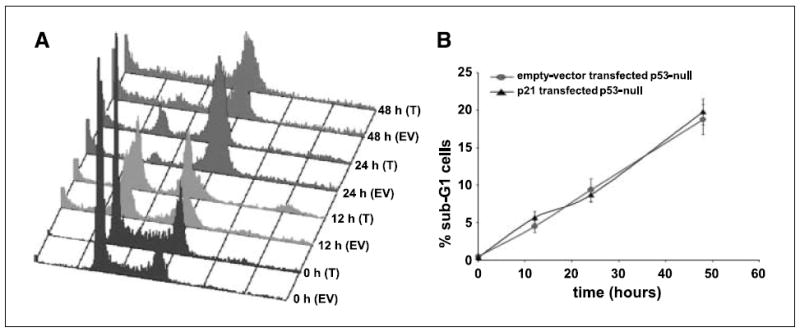
We have previously discovered the naturally occurring antitussive alkaloid noscapine as a tubulin-binding agent that attenuates microtubule dynamics and arrests mammalian cells at mitosis via activation of the c-Jun NH(2)-terminal kinase pathway. It is well established that the p53 protein plays a crucial role in the control of tumor cell response to chemotherapeutic agents and DNA-damaging agents; however, the relationship between p53-driven genes and drug sensitivity remains controversial. In this study, we compared chemosensitivity, cell cycle distribution, and apoptosis on noscapine treatment in four cell lines derived from the colorectal carcinoma HCT116 cells: p53(+/+) (p53-wt), p53(-/-) (p53-null), p21(-/-) (p21-null), and BAX(-/-) (BAX-null). Using these isogenic variants, we investigated the roles of p53, BAX, and p21 in the cellular response to treatment with noscapine. Our results show that noscapine treatment increases the expression of p53 over time in cells with wild-type p53 status. This increase in p53 is associated with an increased apoptotic BAX/Bcl-2 ratio consistent with increased sensitivity of these cells to apoptotic stimuli. Conversely, loss of p53 and p21 alleles had a counter effect on both BAX and Bcl-2 expression and the p53-null and p21-null cells were significantly resistant to the antiproliferative and apoptotic effects of noscapine. All but the p53-null cells displayed p53 protein accumulation in a time-dependent manner on noscapine treatment. Interestingly, despite increased levels of p53, p21-null cells were resistant to apoptosis, suggesting a proapoptotic role of p21 and implying that p53 is a necessary but not sufficient condition for noscapine-mediated apoptosis.
Figures
References
-
- Joshi HC. Microtubule dynamics in living cells. Curr Opin Cell Biol. 1998;10:35–44. - PubMed
-
- Jordan MA, Wendell K, Gardiner S, Derry WB, Copp H, Wilson L. Mitotic block induced in HeLa cells by low concentrations of paclitaxel (Taxol) results in abnormal mitotic exit and apoptotic cell death. Cancer Res. 1996;56:816–25. - PubMed
-
- Gundersen GG, Cook TA. Microtubules and signal transduction. Curr Opin Cell Biol. 1999;11:81–94. - PubMed
-
- Meek DW. Multisite phosphorylation and the integration of stress signals at p53. Cell Signal. 1998;10:159–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
