

A glycogen phosphorylase inhibitor selectively enhances local rates of glucose utilization in brain during sensory stimulation of conscious rats: implications for glycogen turnover
- PMID: 17442042
- PMCID: PMC2822402
- DOI: 10.1111/j.1471-4159.2007.04595.x
A glycogen phosphorylase inhibitor selectively enhances local rates of glucose utilization in brain during sensory stimulation of conscious rats: implications for glycogen turnover
Abstract
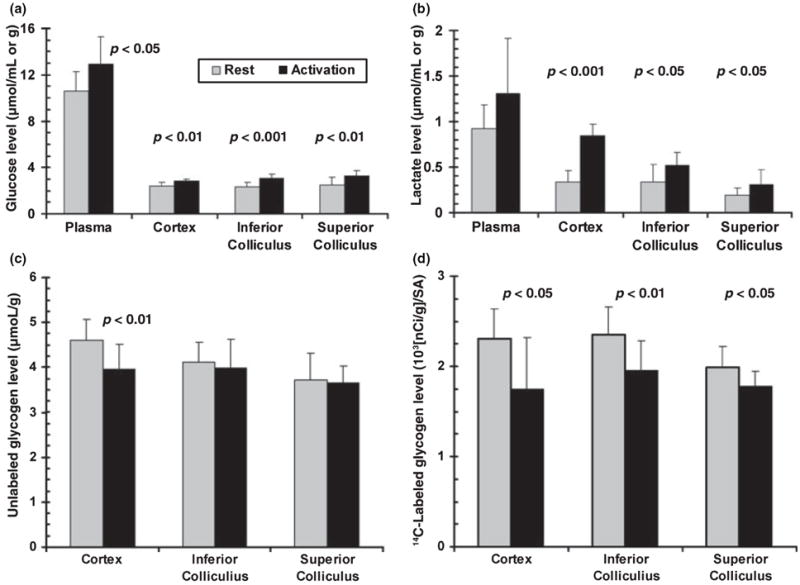
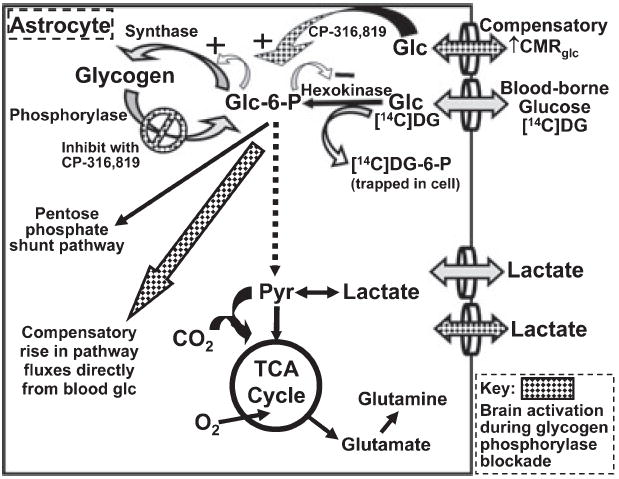
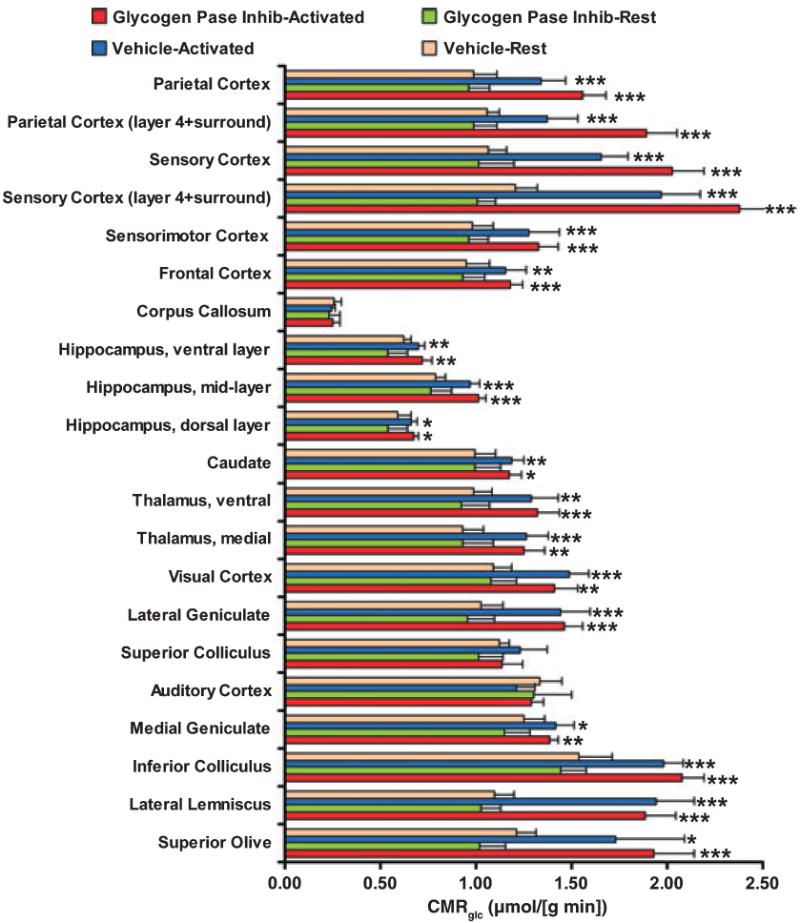
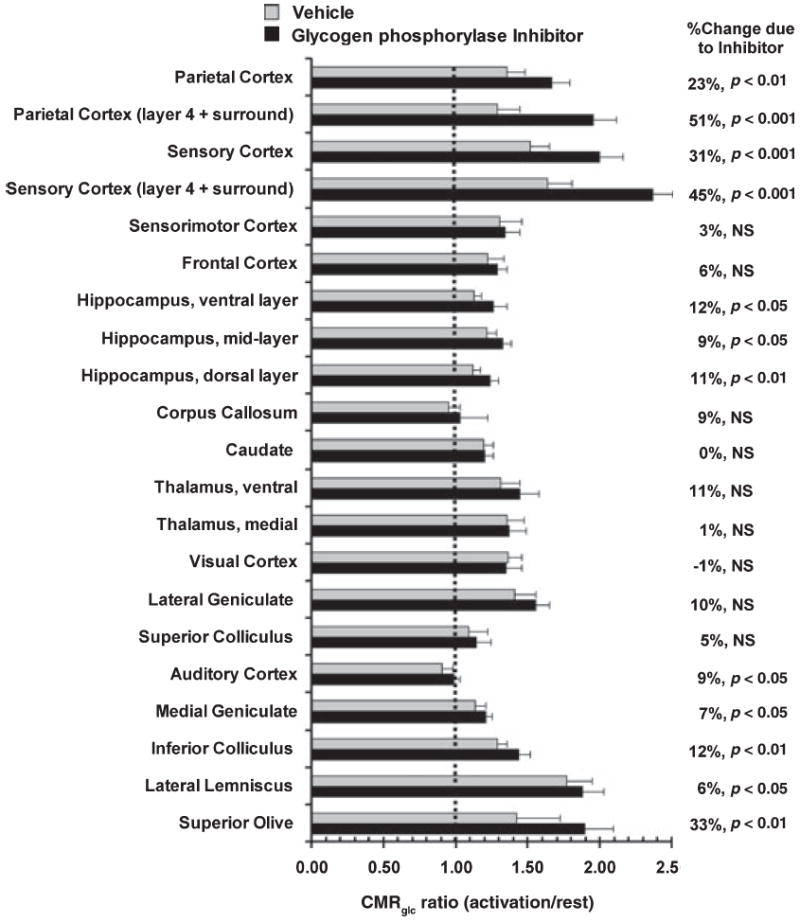
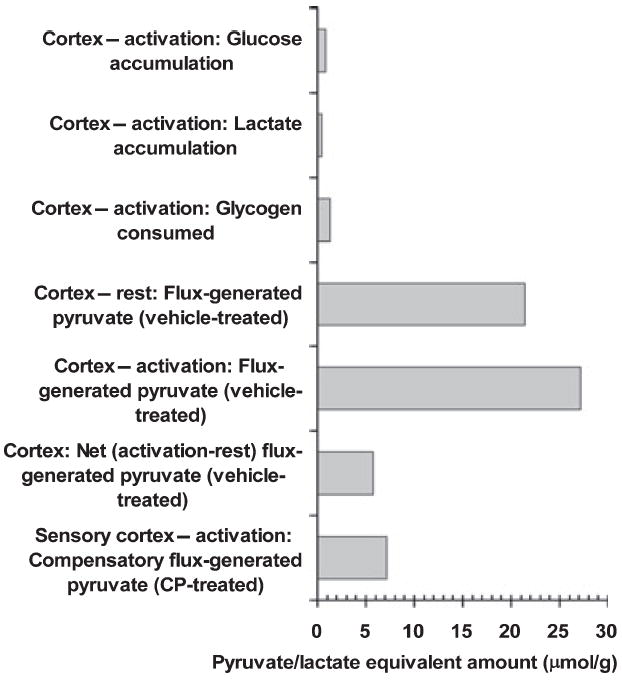
Glycogen is degraded during brain activation but its role and contribution to functional energetics in normal activated brain have not been established. In the present study, glycogen utilization in brain of normal conscious rats during sensory stimulation was assessed by three approaches, change in concentration, release of (14)C from pre-labeled glycogen and compensatory increase in utilization of blood glucose (CMR(glc)) evoked by treatment with a glycogen phosphorylase inhibitor. Glycogen level fell in cortex, (14)C release increased in three structures and inhibitor treatment caused regionally selective compensatory increases in CMR(glc) over and above the activation-induced rise in vehicle-treated rats. The compensatory rise in CMR(glc) was highest in sensory-parietal cortex where it corresponded to about half of the stimulus-induced rise in CMR(glcf) in vehicle-treated rats; this response did not correlate with metabolic rate, stimulus-induced rise in CMR(glc) or sequential station in sensory pathway. Thus, glycogen is an active fuel for specific structures in normal activated brain, not simply an emergency fuel depot and flux-generated pyruvate greatly exceeded net accumulation of lactate or net consumption of glycogen during activation. The metabolic fate of glycogen is unknown, but adding glycogen to the fuel consumed during activation would contribute to a fall in CMR(O2)/CMR(glc) ratio.
Figures
References
-
- Ackermann RF, Lear JL. Glycolysis-induced discordance between glucose metabolic rates measured with radiolabeled fluorodeoxyglucose and glucose. J Cereb Blood Flow Metab. 1989;9:774–785. - PubMed
-
- Beck T, Wree A, Schleicher A. Glucose utilization in rat hippocampus after long-term recovery from ischemia. J Cereb Blood Flow Metab. 1990;10:542–549. - PubMed
-
- sBielajew C, Konkle AT, Merali Z. The effects of chronic mild stress on male Sprague-Dawley and Long Evans rats: I. Biochemical and physiological analyses. Behav Brain Res. 2002;136:583–592. - PubMed
-
- Bielajew C, Konkle AT, Kentner AC, Baker SL, Stewart A, Hutchins AA, Santa-Maria Barbagallo L, Fouriezos G. Strain and gender specific effects in the forced swim test: effects of previous stress exposure. Stress. 2003;6:269–280. - PubMed
-
- Brown AM. Brain glycogen re-awakened. J Neurochem. 2004;89:537–552. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
