

The Hectd1 ubiquitin ligase is required for development of the head mesenchyme and neural tube closure
- PMID: 17442300
- PMCID: PMC2730518
- DOI: 10.1016/j.ydbio.2007.03.018
The Hectd1 ubiquitin ligase is required for development of the head mesenchyme and neural tube closure
Abstract
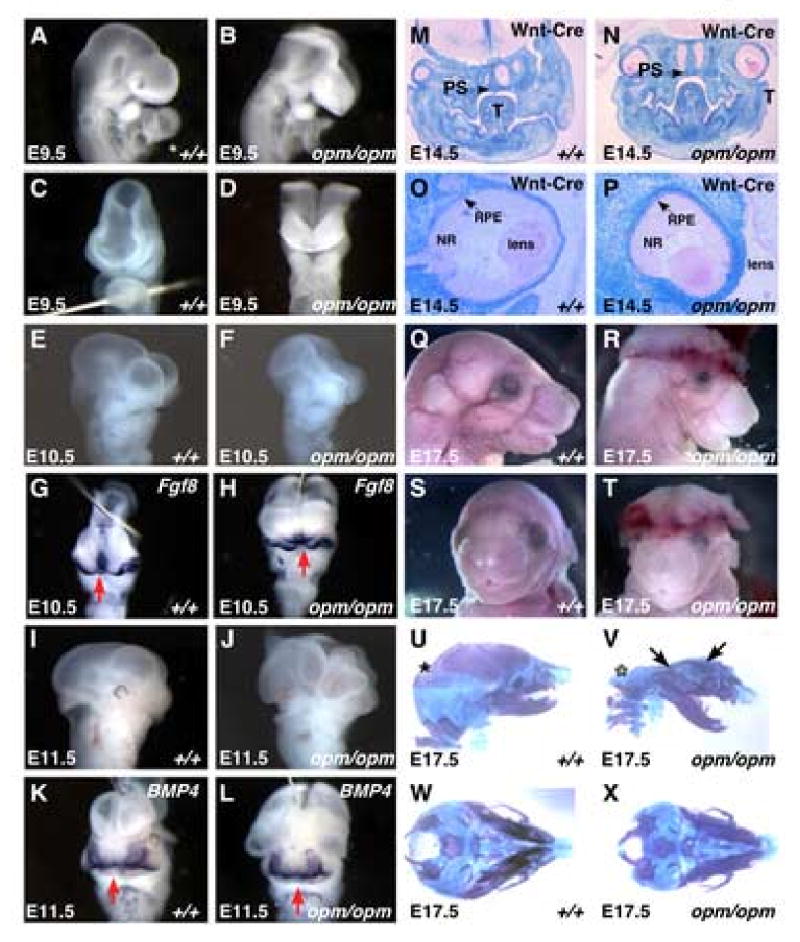
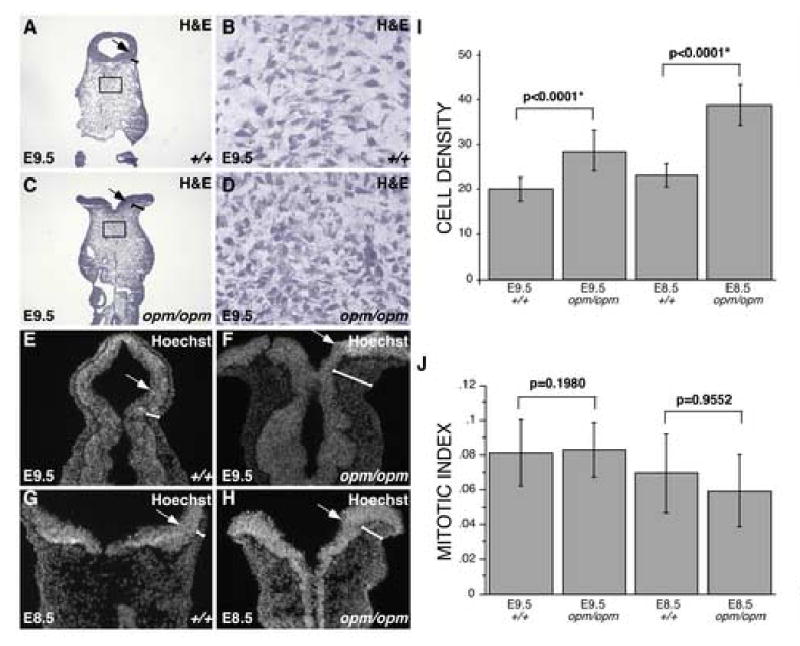
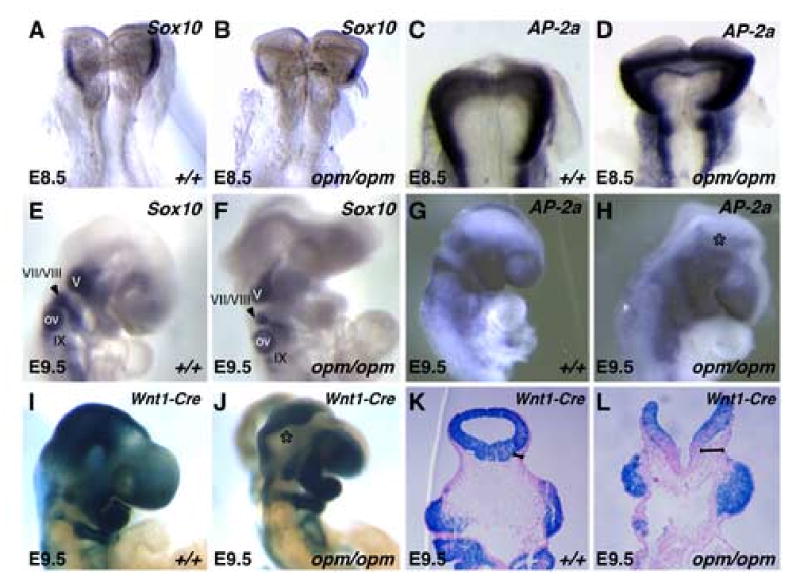
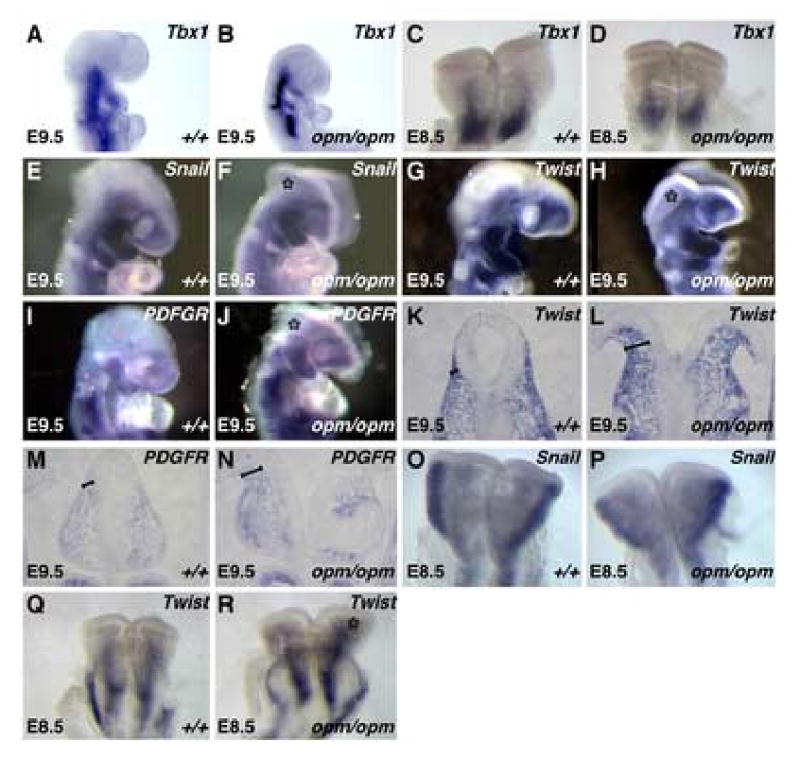
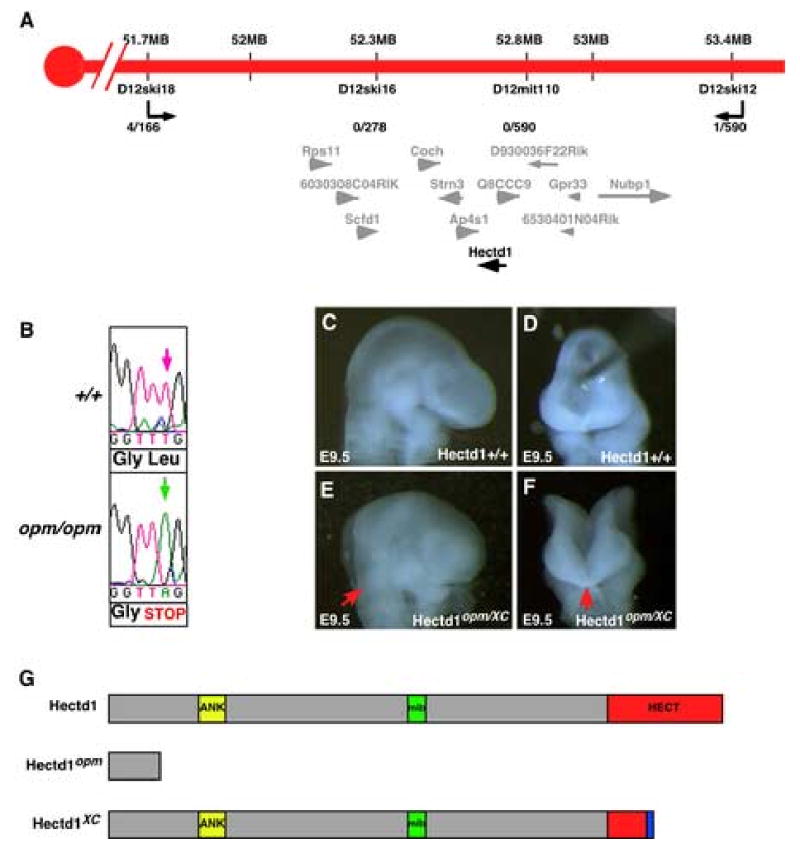
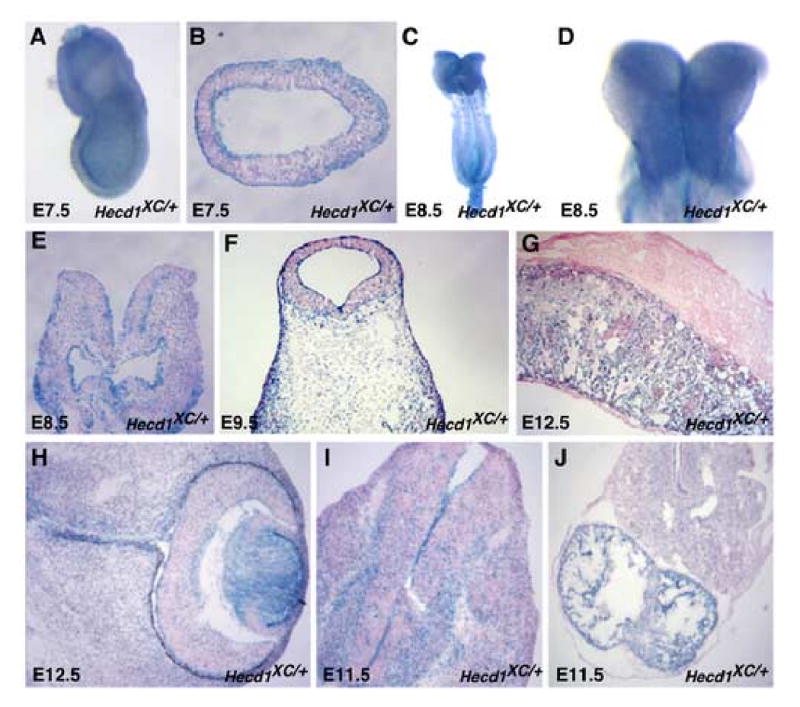
Closure of the cranial neural tube depends on normal development of the head mesenchyme. Homozygous-mutant embryos for the ENU-induced open mind (opm) mutation exhibit exencephaly associated with defects in head mesenchyme development and dorsal-lateral hinge point formation. The head mesenchyme in opm mutant embryos is denser than in wildtype embryos and displays an abnormal cellular organization. Since cells that originate from both the cephalic paraxial mesoderm and the neural crest populate the head mesenchyme, we explored the origin of the abnormal head mesenchyme. opm mutant embryos show apparently normal development of neural crest-derived structures. Furthermore, the abnormal head mesenchyme in opm mutant embryos is not derived from the neural crest, but instead expresses molecular markers of cephalic mesoderm. We also report the identification of the opm mutation in the ubiquitously expressed Hectd1 E3 ubiquitin ligase. Two different Hectd1 alleles cause incompletely penetrant neural tube defects in heterozygous animals, indicating that Hectd1 function is required at a critical threshold for neural tube closure. This low penetrance of neural tube defects in embryos heterozygous for Hectd1 mutations suggests that Hectd1 should be considered as candidate susceptibility gene in human neural tube defects.
Figures
References
-
- Brewer S, Feng W, Huang J, Sullivan S, Williams T. Wnt1-Cre-mediated deletion of AP-2alpha causes multiple neural crest-related defects. Dev Biol. 2004;267:135–52. - PubMed
-
- Caspary T, Anderson KV. Patterning cell types in the dorsal spinal cord: what the mouse mutants say. Nat Rev Neurosci. 2003;4:289–97. - PubMed
-
- Chapman DL, Garvey N, Hancock S, Alexiou M, Agulnik SI, Gibson-Brown JJ, Cebra-Thomas J, Bollag RJ, Silver LM, Papaioannou VE. Expression of the T-box family genes, Tbx1-Tbx5, during early mouse development. Dev Dyn. 1996;206:379–90. - PubMed
-
- Chapman SC, Bernard DJ, Jelen J, Woodruff TK. Properties of inhibin binding to betaglycan, InhBP/p120 and the activin type II receptors. Mol Cell Endocrinol. 2002;196:79–93. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
