

Dietary docosahexaenoic acid and docosapentaenoic acid ameliorate amyloid-beta and tau pathology via a mechanism involving presenilin 1 levels
- PMID: 17442823
- PMCID: PMC6672302
- DOI: 10.1523/JNEUROSCI.0055-07.2007
Dietary docosahexaenoic acid and docosapentaenoic acid ameliorate amyloid-beta and tau pathology via a mechanism involving presenilin 1 levels
Abstract
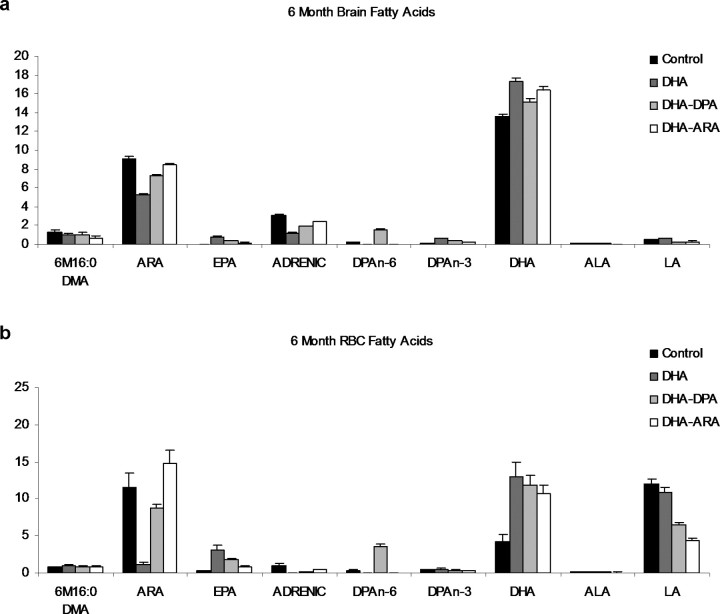
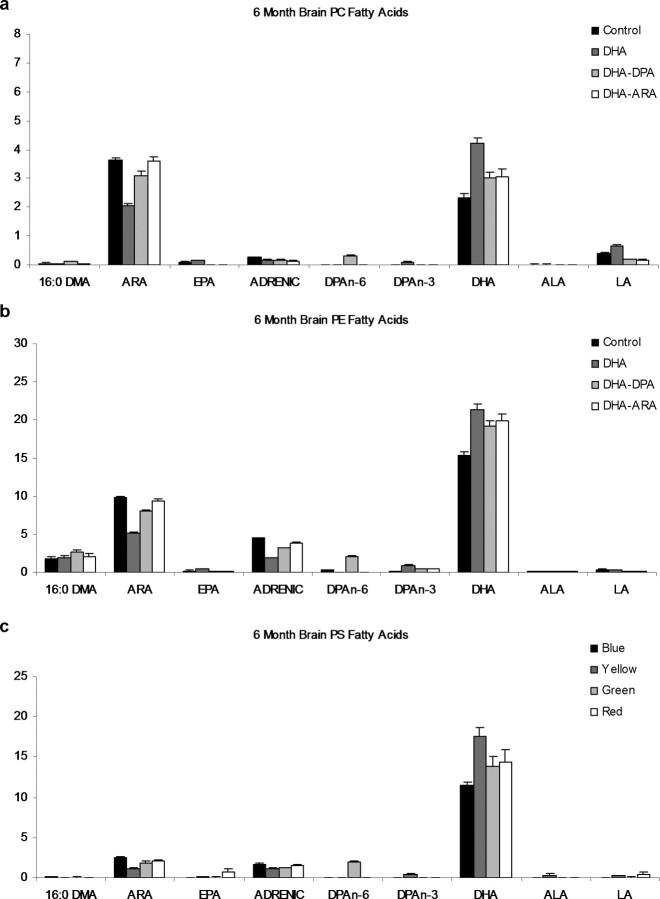
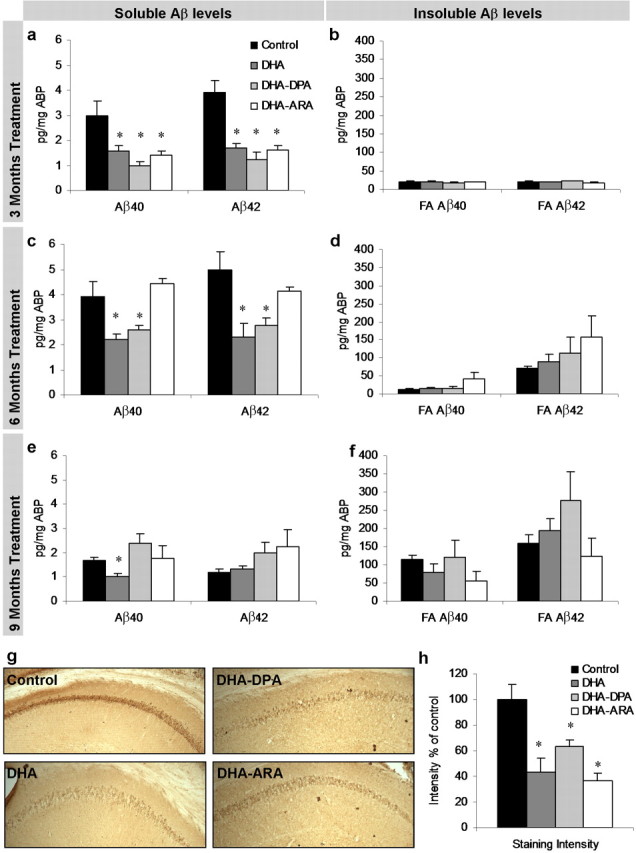
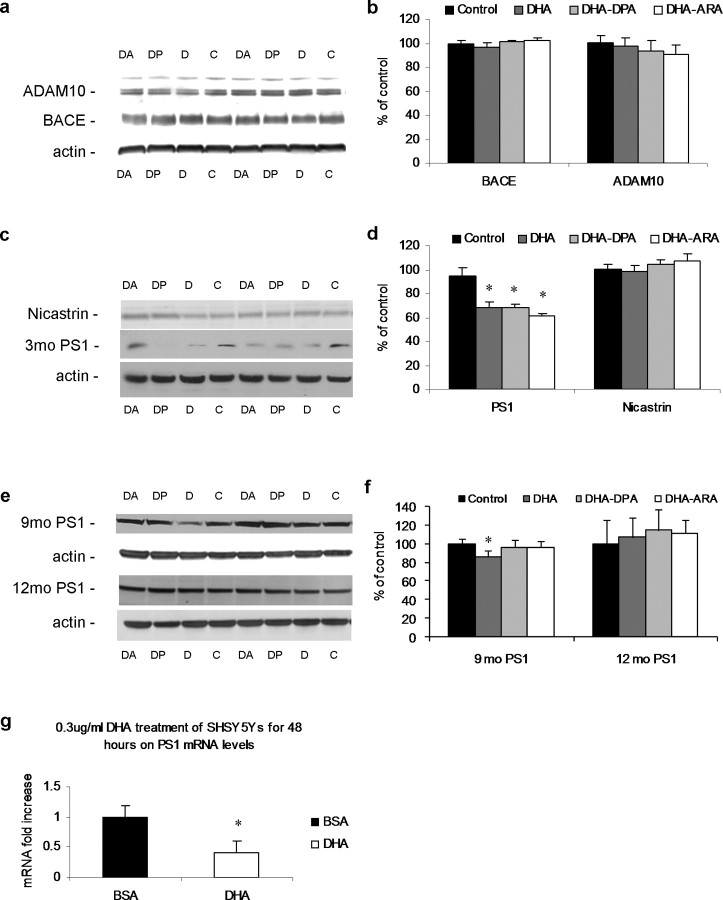
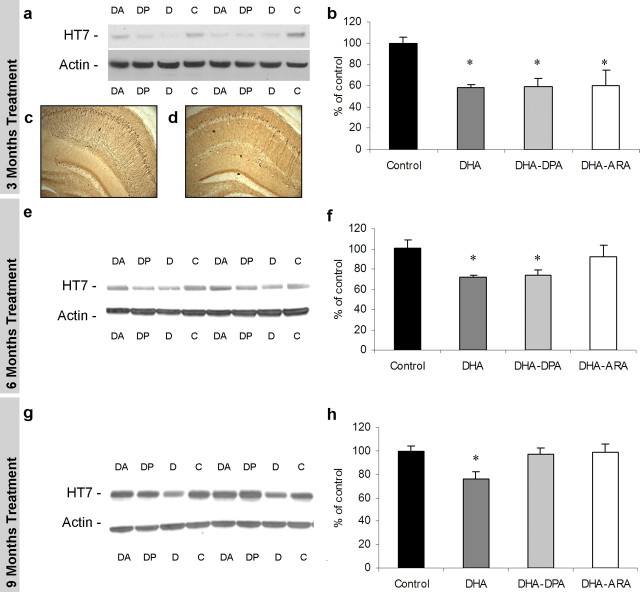
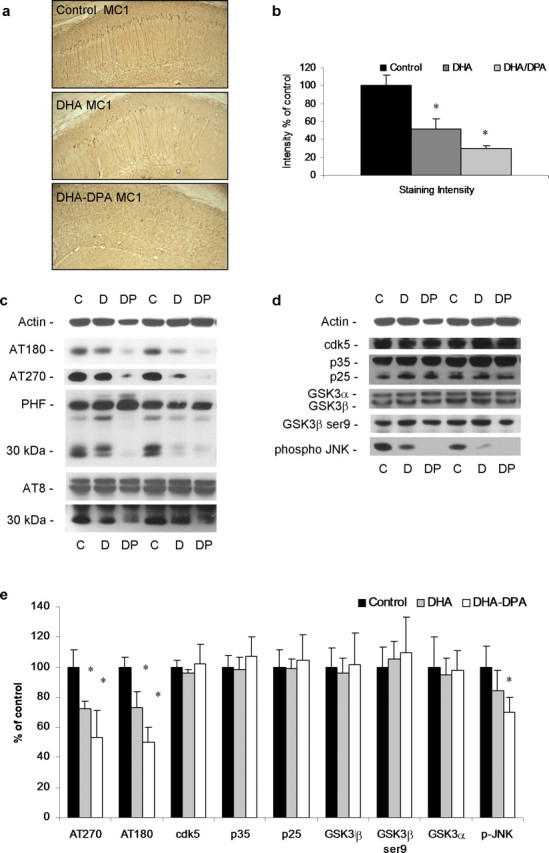
The underlying cause of sporadic Alzheimer disease (AD) is unknown, but a number of environmental and genetic factors are likely to be involved. One environmental factor that is increasingly being recognized as contributing to brain aging is diet, which has evolved markedly over modern history. Here we show that dietary supplementation with docosahexaenoic acid (DHA), an n-3 polyunsaturated fatty acid, in the 3xTg-AD mouse model of AD reduced the intraneuronal accumulation of both amyloid-beta (Abeta) and tau. In contrast, combining DHA with n-6 fatty acids, either arachidonic acid or docosapentaenoic acid (DPAn-6), diminished the efficacy of DHA over a 12 month period. Here we report the novel finding that the mechanism accounting for the reduction in soluble Abeta was attributable to a decrease in steady-state levels of presenilin 1, and not to altered processing of the amyloid precursor protein by either the alpha- or beta-secretase. Furthermore, the presence of DPAn-6 in the diet reduced levels of early-stage phospho-tau epitopes, which correlated with a reduction in phosphorylated c-Jun N-terminal kinase, a putative tau kinase. Collectively, these results suggest that DHA and DPAn-6 supplementations could be a beneficial natural therapy for AD.
Figures

Similar articles
-
High dietary consumption of trans fatty acids decreases brain docosahexaenoic acid but does not alter amyloid-beta and tau pathologies in the 3xTg-AD model of Alzheimer's disease.Neuroscience. 2009 Mar 3;159(1):296-307. doi: 10.1016/j.neuroscience.2008.12.006. Epub 2008 Dec 14. Neuroscience. 2009. PMID: 19135506
-
Endogenous conversion of omega-6 into omega-3 fatty acids improves neuropathology in an animal model of Alzheimer's disease.J Alzheimers Dis. 2011;27(4):853-69. doi: 10.3233/JAD-2011-111010. J Alzheimers Dis. 2011. PMID: 21914946
-
Differential impact of eicosapentaenoic acid and docosahexaenoic acid in an animal model of Alzheimer's disease.J Lipid Res. 2024 Dec;65(12):100682. doi: 10.1016/j.jlr.2024.100682. Epub 2024 Oct 28. J Lipid Res. 2024. PMID: 39490923 Free PMC article.
-
Docosahexaenoic acid protects from amyloid and dendritic pathology in an Alzheimer's disease mouse model.Nutr Health. 2006;18(3):249-59. doi: 10.1177/026010600601800307. Nutr Health. 2006. PMID: 17180870 Review.
-
The Impact of Docosahexaenoic Acid on Alzheimer's Disease: Is There a Role of the Blood-Brain Barrier?Curr Clin Pharmacol. 2015;10(3):222-41. doi: 10.2174/157488471003150820151532. Curr Clin Pharmacol. 2015. PMID: 26338174 Review.
Cited by
-
Precision Nutrition for Alzheimer's Prevention in ApoE4 Carriers.Nutrients. 2021 Apr 19;13(4):1362. doi: 10.3390/nu13041362. Nutrients. 2021. PMID: 33921683 Free PMC article. Review.
-
The effect of dietary supplementation on brain-derived neurotrophic factor and cognitive functioning in Alzheimer's dementia.J Clin Transl Res. 2017 Dec 3;3(3):337-343. eCollection 2018 Jan 15. J Clin Transl Res. 2017. PMID: 30895275 Free PMC article.
-
Omega-3 Fatty Acid-Type Docosahexaenoic Acid Protects against Aβ-Mediated Mitochondrial Deficits and Pathomechanisms in Alzheimer's Disease-Related Animal Model.Int J Mol Sci. 2020 May 29;21(11):3879. doi: 10.3390/ijms21113879. Int J Mol Sci. 2020. PMID: 32486013 Free PMC article.
-
Beta-amyloid oligomers induce phosphorylation of tau and inactivation of insulin receptor substrate via c-Jun N-terminal kinase signaling: suppression by omega-3 fatty acids and curcumin.J Neurosci. 2009 Jul 15;29(28):9078-89. doi: 10.1523/JNEUROSCI.1071-09.2009. J Neurosci. 2009. PMID: 19605645 Free PMC article.
-
Role of early life exposure and environment on neurodegeneration: implications on brain disorders.Transl Neurodegener. 2014 Apr 29;3:9. doi: 10.1186/2047-9158-3-9. eCollection 2014. Transl Neurodegener. 2014. PMID: 24847438 Free PMC article. Review.
References
-
- Agiostratidou G, Muros RM, Shioi J, Marambaud P, Robakis NK. The cytoplasmic sequence of E-cadherin promotes non-amyloidogenic degradation of A beta precursors. J Neurochem. 2006;96:1182–1188. - PubMed
-
- Ansari KA, Shoeman DW. Arachidonic and docosahexanoic acid content of bovine brain myelin: implications for the pathogenesis of multiple sclerosis. Neurochem Res. 1990;15:7–11. - PubMed
-
- Arai T, Guo JP, McGeer PL. Proteolysis of non-phosphorylated and phosphorylated tau by thrombin. J Biol Chem. 2005;280:5145–5153. - PubMed
-
- Bazan NG, Scott BL. Dietary omega-3 fatty acids and accumulation of docosahexaenoic acid in rod photoreceptor cells of the retina and at synapses. Ups J Med Sci Suppl. 1990;48:97–107. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous