

Site-specific S-glutathiolation of mitochondrial NADH ubiquinone reductase
- PMID: 17444656
- PMCID: PMC2527596
- DOI: 10.1021/bi602580c
Site-specific S-glutathiolation of mitochondrial NADH ubiquinone reductase
Abstract
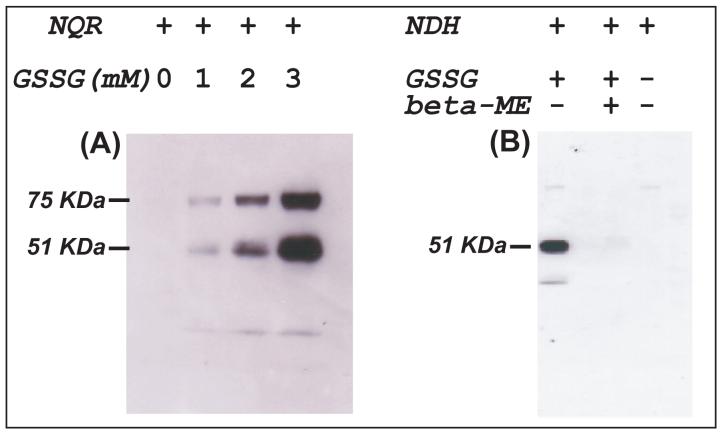
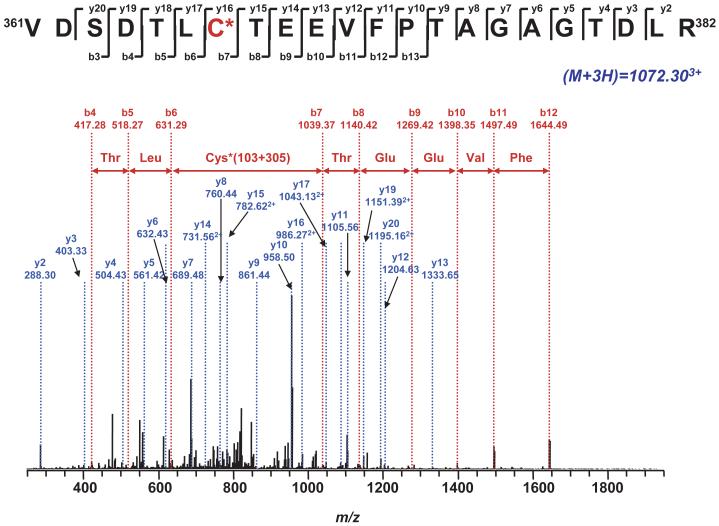
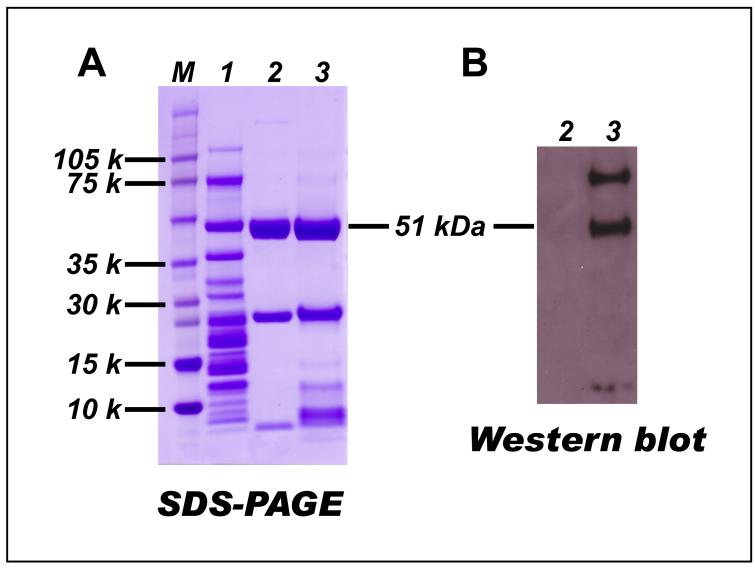
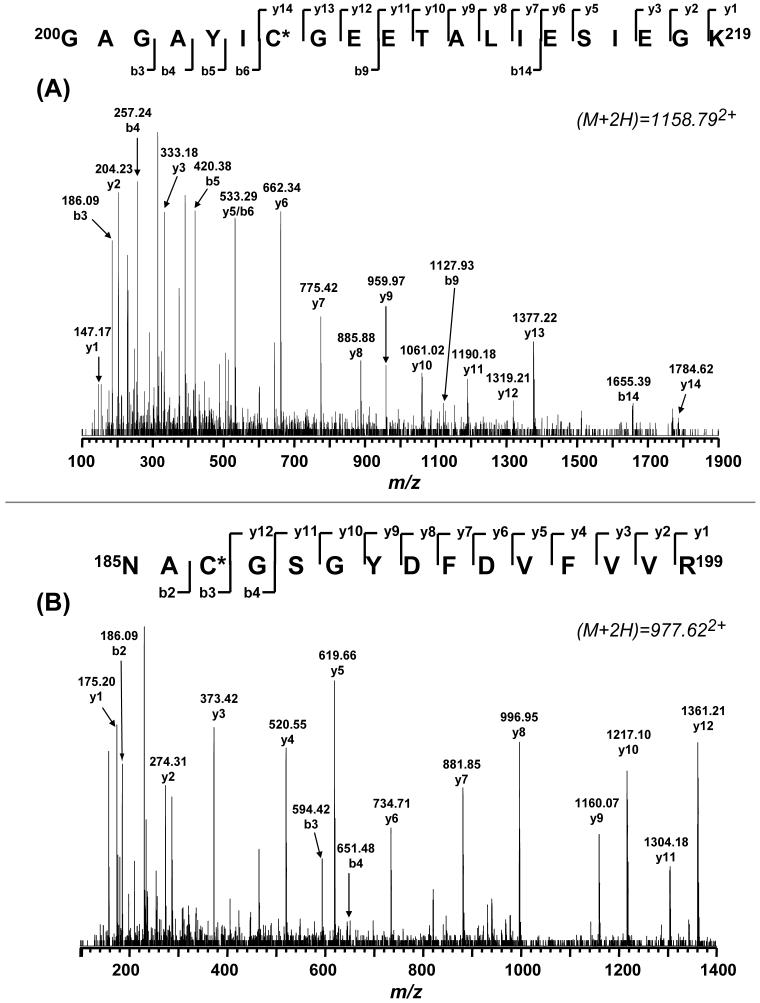
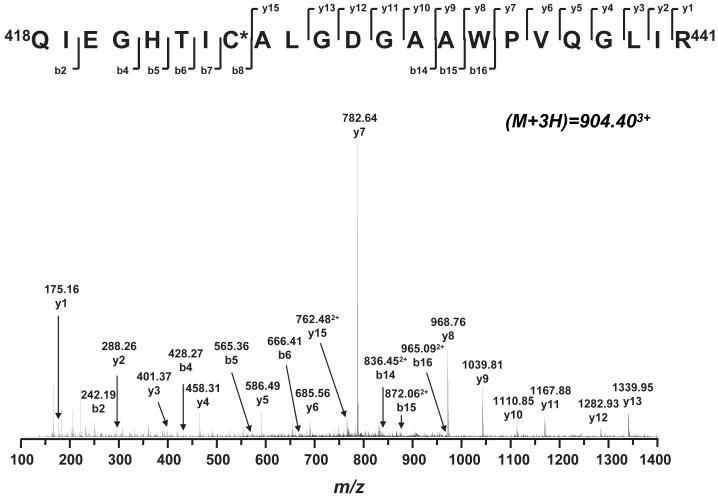
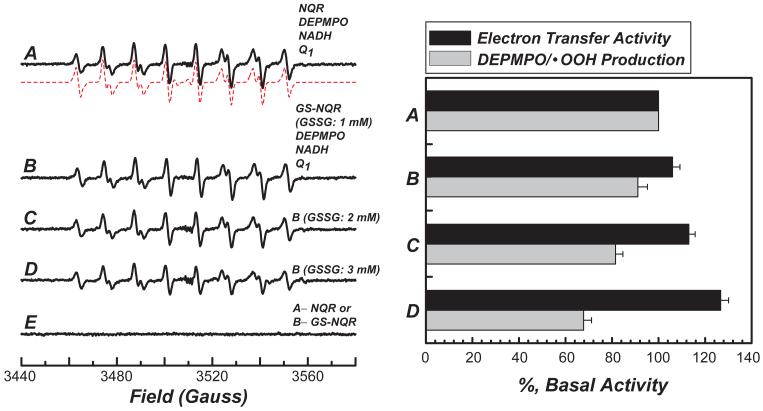
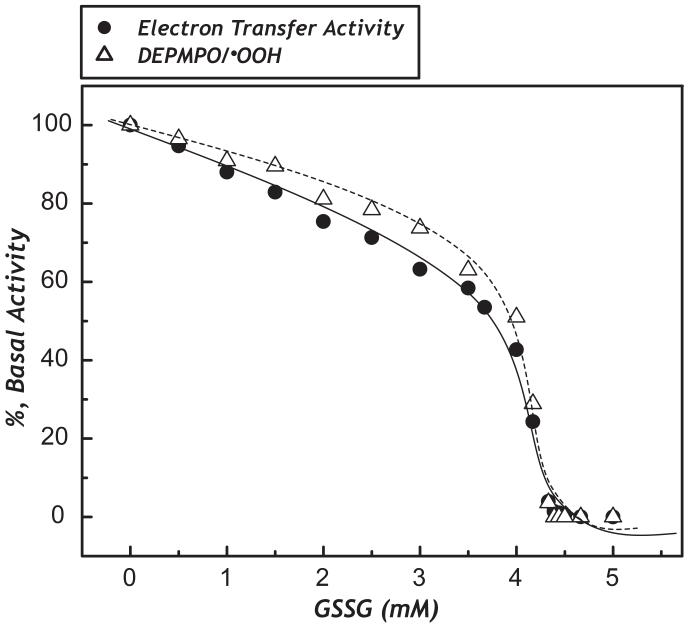
The generation of reactive oxygen species in mitochondria acts as a redox signal in triggering cellular events such as apoptosis, proliferation, and senescence. Overproduction of superoxide (O2*-) and O2*--derived oxidants changes the redox status of the mitochondrial GSH pool. An electron transport protein, mitochondrial complex I, is the major host of reactive/regulatory protein thiols. An important response of protein thiols to oxidative stress is to reversibly form protein mixed disulfide via S-glutathiolation. Exposure of complex I to oxidized GSH, GSSG, resulted in specific S-glutathiolation at the 51 kDa and 75 kDa subunits (Beer et al. (2004) J. Biol. Chem. 279, 47939-47951). Here, to investigate the molecular mechanism of S-glutathiolation of complex I, we prepared isolated bovine complex I under nonreducing conditions and employed the techniques of mass spectrometry and EPR spin trapping for analysis. LC/MS/MS analysis of tryptic digests of the 51 kDa and 75 kDa polypeptides from glutathiolated complex I (GS-NQR) revealed that two specific cysteines (C206 and C187) of the 51 kDa subunit and one specific cysteine (C367) of the 75 kDa subunit were involved in redox modifications with GS binding. The electron transfer activity (ETA) of GS-NQR in catalyzing NADH oxidation by Q1 was significantly enhanced. However, O2*- generation activity (SGA) mediated by GS-NQR suffered a mild loss as measured by EPR spin trapping, suggesting the protective role of S-glutathiolation in the intact complex I. Exposure of NADH dehydrogenase (NDH), the flavin subcomplex of complex I, to GSSG resulted in specific S-glutathiolation on the 51 kDa subunit. Both ETA and SGA of S-glutathiolated NDH (GS-NDH) decreased in parallel as the dosage of GSSG increased. LC/MS/MS analysis of a tryptic digest of the 51 kDa subunit from GS-NDH revealed that C206, C187, and C425 were glutathiolated. C425 of the 51 kDa subunit is a ligand residue of the 4Fe-4S N3 center, suggesting that destruction of 4Fe-4S is the major mechanism involved in the inhibition of NDH. The result also implies that S-glutathiolation of the 75 kDa subunit may play a role in protecting the 4Fe-4S cluster of the 51 kDa subunit from redox modification when complex I is exposed to redox change in the GSH pool.
Figures


References
-
- Hirst J, Carroll J, Fearnley IM, Shannon RJ, Walker JE. The nuclear encoded subunits of complex I from bovine heart mitochondria. Biochim Biophys Acta. 2003;1604:135–50. - PubMed
-
- Carroll J, Fearnley IM, Shannon RJ, Hirst J, Walker JE. Analysis of the subunit composition of complex I from bovine heart mitochondria. Mol Cell Proteomics. 2003;2:117–26. - PubMed
-
- Walker JE. The NADH:ubiquinone oxidoreductase (complex I) of respiratory chains. Q Rev Biophys. 1992;25:253–324. - PubMed
-
- Galkin A, Brandt U. Superoxide radical formation by pure complex I (NADH:ubiquinone oxidoreductase) from Yarrowia lipolytica. J Biol Chem. 2005;280:30129–35. - PubMed
-
- Kudin AP, Bimpong-Buta NY, Vielhaber S, Elger CE, Kunz WS. Characterization of superoxide-producing sites in isolated brain mitochondria. J Biol Chem. 2004;279:4127–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
