

Axonal regeneration and development of de novo axons from distal dendrites of adult feline commissural interneurons after a proximal axotomy
- PMID: 17447249
- PMCID: PMC2930906
- DOI: 10.1002/cne.21362
Axonal regeneration and development of de novo axons from distal dendrites of adult feline commissural interneurons after a proximal axotomy
Abstract
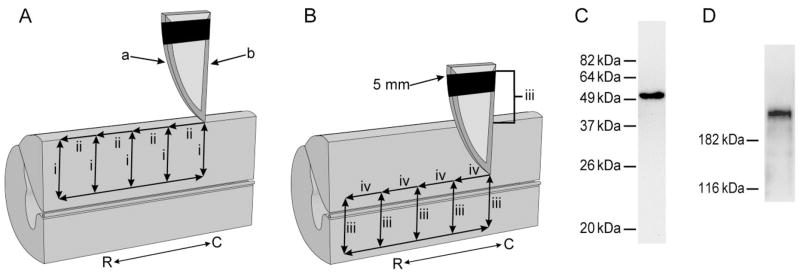
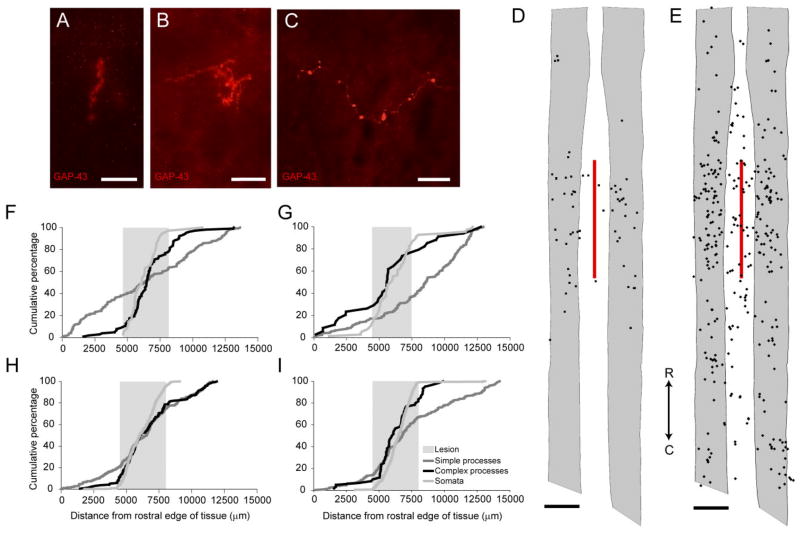
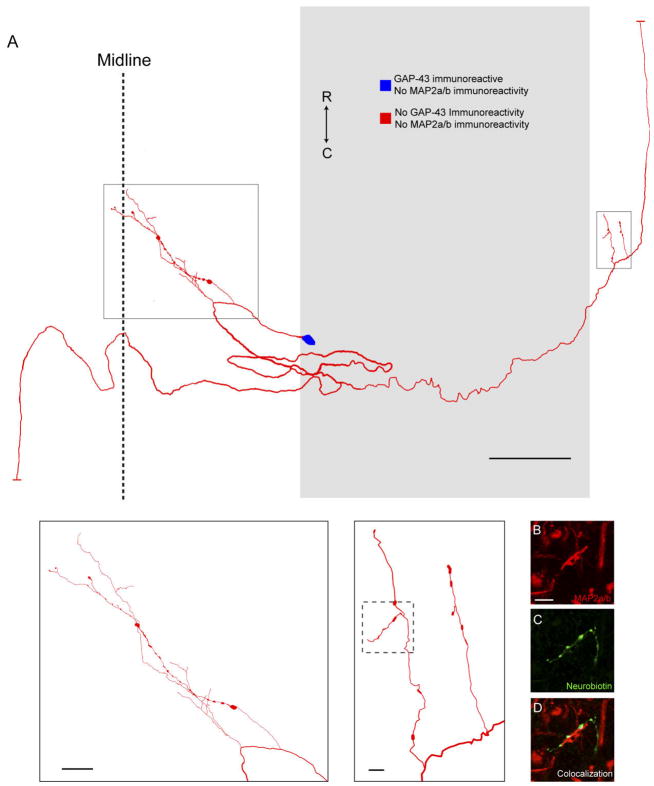
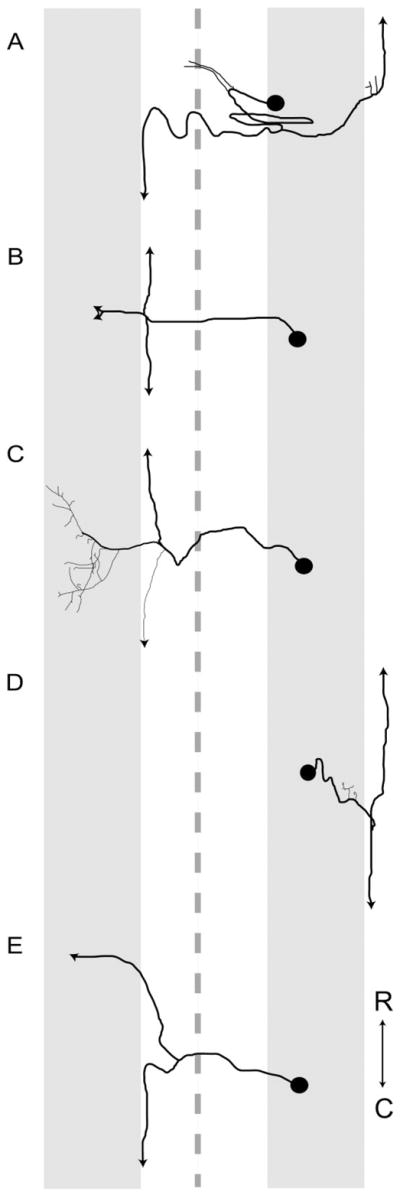
Following proximal axotomy, several types of neurons sprout de novo axons from distal dendrites. These processes may represent a means of forming new circuits following spinal cord injury. However, it is not know whether mammalian spinal interneurons, axotomized as a result of a spinal cord injury, develop de novo axons. Our goal was to determine whether spinal commissural interneurons (CINs), axotomized by 3-4-mm midsagittal transection at C3, form de novo axons from distal dendrites. All experiments were performed on adult cats. CINs in C3 were stained with extracellular injections of Neurobiotin at 4-5 weeks post injury. The somata of axotomized CINs were identified by the presence of immunoreactivity for the axonal growth-associated protein-43 (GAP-43). Nearly half of the CINs had de novo axons that emerged from distal dendrites. These axons lacked immunoreactivity for the dendritic protein, microtubule-associated protein2a/b (MAP2a/b); some had GAP-43-immunoreactive terminals; and nearly all had morphological features typical of axons. Dendrites of other CINs did not give rise to de novo axons. These CINs did, however, each have a long axon-like process (L-ALP) that projected directly from the soma or a very proximal dendrite. L-ALPs were devoid of MAP2a/b immunoreactivity. Some of these L-ALPs projected through the lesion and formed bouton-like swellings. These results suggest that proximally axotomized spinal interneurons have the potential to form new connections via de novo axons that emerge from distal dendrites. Others may be capable of regeneration of their original axon.
(c) 2007 Wiley-Liss, Inc.
Figures







Similar articles
-
The temporal sequence of morphological and molecular changes in axotomized feline motoneurons leading to the formation of axons from the ends of dendrites.J Comp Neurol. 2004 Jan 6;468(2):233-50. doi: 10.1002/cne.10966. J Comp Neurol. 2004. PMID: 14648682
-
Alterations to neuronal polarity following permanent axotomy: a quantitative analysis of changes to MAP2a/b and GAP-43 distributions in axotomized motoneurons in the adult cat.J Comp Neurol. 2002 Sep 2;450(4):318-33. doi: 10.1002/cne.10324. J Comp Neurol. 2002. PMID: 12209846
-
Emergence of axons from distal dendrites of adult mammalian neurons following a permanent axotomy.Eur J Neurosci. 2001 Mar;13(6):1166-76. doi: 10.1046/j.0953-816x.2001.1490.x. Eur J Neurosci. 2001. PMID: 11285014
-
Control of neuron shape during development and regeneration.Neurochem Pathol. 1986 Dec;5(3):331-43. doi: 10.1007/BF02842942. Neurochem Pathol. 1986. PMID: 3306477 Review.
-
Determinants of Axon Growth, Plasticity, and Regeneration in the Context of Spinal Cord Injury.Am J Pathol. 2018 Jan;188(1):53-62. doi: 10.1016/j.ajpath.2017.09.005. Epub 2017 Oct 10. Am J Pathol. 2018. PMID: 29030051 Free PMC article. Review.
Cited by
-
Mechanisms of Axon Growth and Regeneration: Moving between Development and Disease.J Neurosci. 2022 Nov 9;42(45):8393-8405. doi: 10.1523/JNEUROSCI.1131-22.2022. J Neurosci. 2022. PMID: 36351827 Free PMC article.
-
Aberrant information transfer interferes with functional axon regeneration.Elife. 2018 Oct 29;7:e38829. doi: 10.7554/eLife.38829. Elife. 2018. PMID: 30371349 Free PMC article.
-
Anatomical Plasticity of Rostrally Terminating Axons as a Possible Bridging Substrate across a Spinal Injury.J Neurotrauma. 2020 Mar 15;37(6):877-888. doi: 10.1089/neu.2018.6193. Epub 2019 Dec 23. J Neurotrauma. 2020. PMID: 31774025 Free PMC article.
-
Proteins recruited to exosomes by tau overexpression implicate novel cellular mechanisms linking tau secretion with Alzheimer's disease.J Alzheimers Dis. 2014;40 Suppl 1(Suppl 1):S47-70. doi: 10.3233/JAD-132135. J Alzheimers Dis. 2014. PMID: 24718102 Free PMC article.
-
Peripheral facial nerve axotomy in mice causes sprouting of motor axons into perineuronal central white matter: time course and molecular characterization.J Comp Neurol. 2010 Mar 1;518(5):699-721. doi: 10.1002/cne.22240. J Comp Neurol. 2010. PMID: 20034058 Free PMC article.
References
-
- Bareyre FM, Kerschensteiner M, Raineteau O, Mettenleiter TC, Weinmann O, Schwab ME. The injured spinal cord spontaneously forms a new intraspinal circuit in adult rats. Nat Neurosci. 2004;7:269–277. - PubMed
-
- Bolton PS, Goto T, Wilson VJ. Commissural neurons in the cat upper cervical spinal cord. Neuroreport. 1991;2:743–746. - PubMed
-
- Cho EY, So KF. Characterization of the sprouting response of axon-like processes from retinal ganglion cells after axotomy in adult hamsters: a model using intravitreal implantation of a peripheral nerve. J Neurocytol. 1992;21:589–603. - PubMed
-
- Coblentz FE, Radeke MJ, Lewis GP, Fisher SK. Evidence that ganglion cells react to retinal detachment. Exp Eye Res. 2003;76:333–342. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
