

barren inflorescence2 Encodes a co-ortholog of the PINOID serine/threonine kinase and is required for organogenesis during inflorescence and vegetative development in maize
- PMID: 17449648
- PMCID: PMC1914211
- DOI: 10.1104/pp.107.098558
barren inflorescence2 Encodes a co-ortholog of the PINOID serine/threonine kinase and is required for organogenesis during inflorescence and vegetative development in maize
Abstract
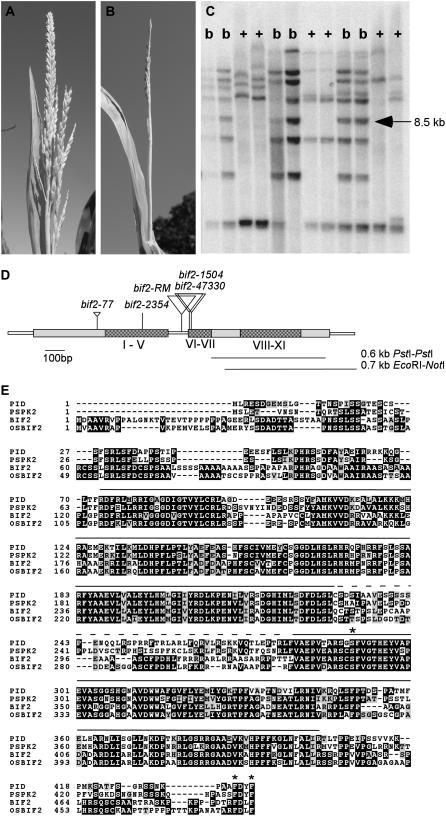
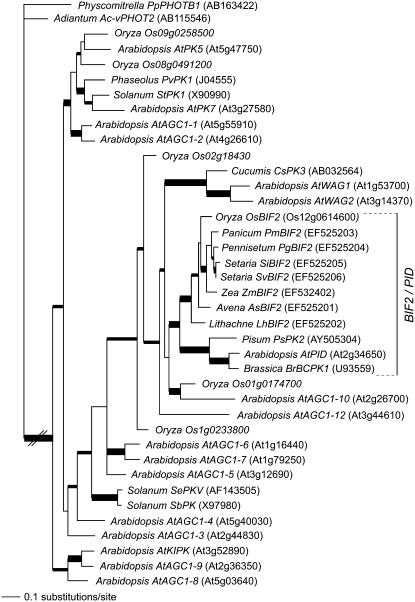
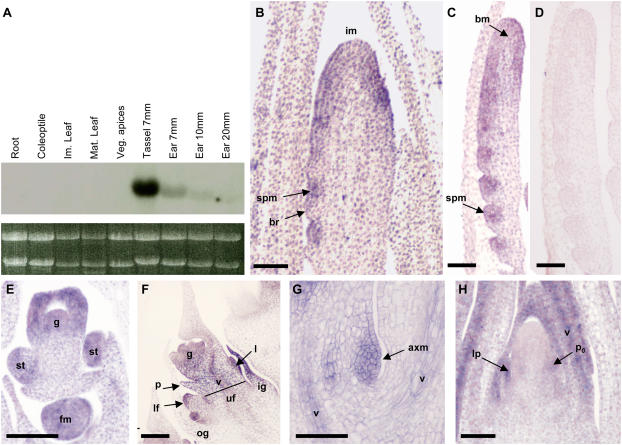
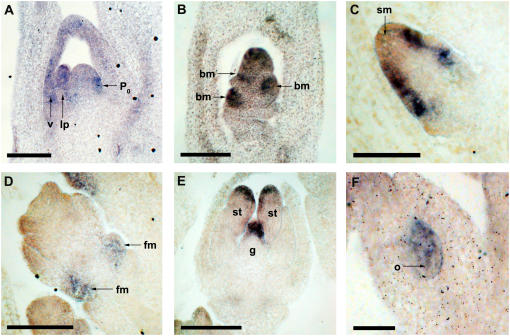
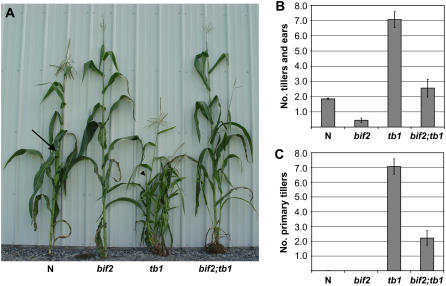
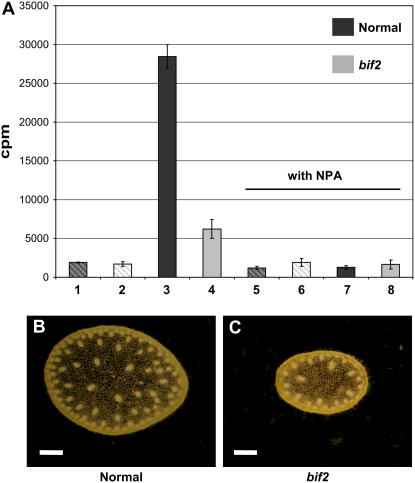
Organogenesis in plants is controlled by meristems. Axillary meristems, which give rise to branches and flowers, play a critical role in plant architecture and reproduction. Maize (Zea mays) and rice (Oryza sativa) have additional types of axillary meristems in the inflorescence compared to Arabidopsis (Arabidopsis thaliana) and thus provide an excellent model system to study axillary meristem initiation. Previously, we characterized the barren inflorescence2 (bif2) mutant in maize and showed that bif2 plays a key role in axillary meristem and lateral primordia initiation in the inflorescence. In this article, we cloned bif2 by transposon tagging. Isolation of bif2-like genes from seven other grasses, along with phylogenetic analysis, showed that bif2 is a co-ortholog of PINOID (PID), which regulates auxin transport in Arabidopsis. Expression analysis showed that bif2 is expressed in all axillary meristems and lateral primordia during inflorescence and vegetative development in maize and rice. Further phenotypic analysis of bif2 mutants in maize illustrates additional roles of bif2 during vegetative development. We propose that bif2/PID sequence and expression are conserved between grasses and Arabidopsis, attesting to the important role they play in development. We provide further support that bif2, and by analogy PID, is required for initiation of both axillary meristems and lateral primordia.
Figures
References
-
- Babb S, Muehlbauer GJ (2003) Genetic and morphological characterization of the barley uniculm2 (cul2) mutant. Theor Appl Genet 106 846–857 - PubMed
-
- Bai F, Watson JC, Walling J, Weeden N, Santner AA, DeMason DA (2005) Molecular characterization and expression of PsPK2, a PINOID-like gene from pea (Pisum sativum). Plant Sci 168 1281–1291
-
- Benjamins R, Quint A, Weijers D, Hooykaas P, Offringa R (2001) The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport. Development 128 4057–4067 - PubMed
-
- Bennett SRM, Alvarez J, Bossinger G, Smyth DR (1995) Morphogenesis in pinoid mutants of Arabidopsis thaliana. Plant J 8 505–520
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
