

doi: 10.1111/j.1574-6968.2007.00722.x.
Epub 2007 Apr 20.
Porphyromonas gingivalis short fimbriae are regulated by a FimS/FimR two-component system
Affiliations
- PMID: 17451448
- PMCID: PMC1974823
- DOI: 10.1111/j.1574-6968.2007.00722.x
Item in Clipboard
Porphyromonas gingivalis short fimbriae are regulated by a FimS/FimR two-component system
FEMS Microbiol Lett.
2007 Jun.
Abstract
Porphyromonas gingivalis possesses two distinct fimbriae. The long (FimA) fimbriae have been extensively studied. Expression of the fimA gene is tightly controlled by a two-component system (FimS/FimR) through a cascade regulation. The short (Mfa1) fimbriae are less understood. The authors have recently demonstrated that both fimbriae are required for formation of P. gingivalis biofilms. Here, the novel finding that FimR, a member of the two-component regulatory system, is a transcriptional activator of the mfa1 gene is promoted. Unlike the regulatory mechanism of FimA by FimR, this regulation of the mfa1 gene is accomplished by FimR directly binding to the promoter region of mfa1.
Figures
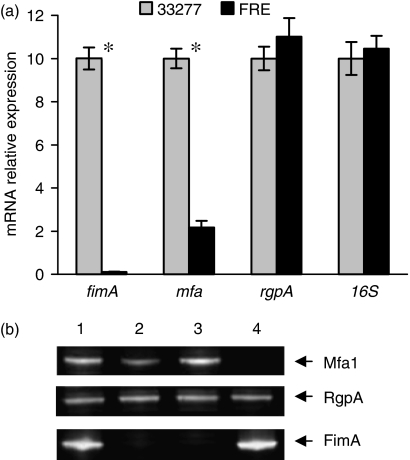
Expression of fimbrial genes in Porphyromonas gingivalis. (a) Fold changes in gene transcription in wild type strain 33277 and the fimR− mutant were measured by real-time PCR. Primers used for each gene are shown in Table 2. Results shown are means and SDs from triplicate experiments. Fold differences were calculated using the relative comparison method. Significant differences (P<0.001 by t test) are labeled with asterisks. (b) Western blot analyses show the expression of Mfa1, RgpA, and FimA in wild type 33 277 (lane 1), the fimR− mutant FRE (lane 2), the fimA− mutant FAT (lane 3), and the mfa1− mutant MFAE (lane 4).

Determination of the transcriptional start site of mfa1. (a) Four different mfa1 sense-strand primers (lanes 2–5, primers MfaTSR2, MfaTSR1, MfaTSR4, MfaTSR3, respectively) were used with 5′ RLM-RACE primers to determine the approximate transcriptional start for mfa1. The PCR products were visualized on a 1.5% agarose TAE gel containing ethidium bromide. Molecular weight standards (indicated in base pairs) are in lane 1. (b) DNA sequence of the mfa1 promoter region. The transcriptional start site A (+1) and the potential start codon ATG are bolded. The primers used for RLM-RACE and RT-PCR are underlined. (c) RT-PCR analysis. Lane 1, 1 kb ladder marker; lane 2 is RT-PCR with MfaTSF3 (from −139 to −121) and MfaTSR5 (from +805 to +824); lane 3 is RT-PCR with MfaTSF2 (from −66 to −47) and MfaTSR5 (from +805 to +824); lane 4 is RT-PCR MfaTSF1 (from +6 to +25) and MfaTSR5 (from +805 to +824).
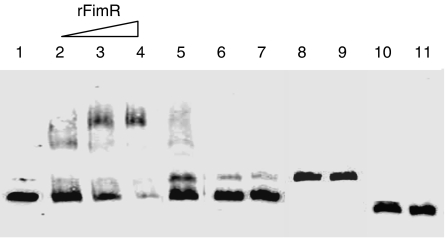
Nucleic acid binding properties of FimR. Electrophoretic mobility shift assays were performed using the biotin-labeled DNA probes. Lane 1, the biotin-labeled mfa1 promoter region (156 bp) alone; lane 2–4, mobility of the biotin-labeled mfa1 promoter region was noted in the presence of increasing amounts of rFimR protein (10, 20, and 40 pmol μL−1, respectively), as indicated on the top; lane 5, the biotin-labeled mfa1 promoter region, rfimR 40 pmol μL−1, and 100-fold excess of unlabeled mfa1 probe; lane 6 is the biotin-labeled mfa1 promoter region alone; lane 7 is the biotin-labeled mfa1 promoter region, and rHGP44 40 pmol μL−1; lane 8 is the biotin-labeled fimA probe (176 bp) alone; lane 9 is the biotin-labeled fimA probe (176 bp), and rFimR 40 pmol μL−1; lane 10 is the biotin-labeled mfa1 coding region (121 bp) alone; lane 11 is the biotin-labeled mfa1 coding region (121 bp) and rFimR 40 pmol μL−1.
References
-
- Amano A, Nakagawa I, Okahashi N, Hamada N. Variations of Porphyromonas gingivalis fimbriae in relation to microbial pathogenesis. J Periodontal Res. 2004;39:136–142. - PubMed
-
- Boggess KA, Madianos PN, Preisser JS, Moise KJ, Jr, Offenbacher S. Chronic maternal and fetal Porphyromonas gingivalis exposure during pregnancy in rabbits. Am J Obstet Gynecol. 2005;192:554–557. - PubMed
-
- Brodala N, Merricks EP, Bellinger DA, et al. Porphyromonas gingivalis bacteremia induces coronary and aortic atherosclerosis in normocholesterolemic and hypercholesterolemic pigs. Arterioscler Thromb Vasc Biol. 2005;25:1446–1451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
