

Linear derivatives of Saccharomyces cerevisiae chromosome III can be maintained in the absence of autonomously replicating sequence elements
- PMID: 17452442
- PMCID: PMC1951491
- DOI: 10.1128/MCB.01246-06
Linear derivatives of Saccharomyces cerevisiae chromosome III can be maintained in the absence of autonomously replicating sequence elements
Abstract
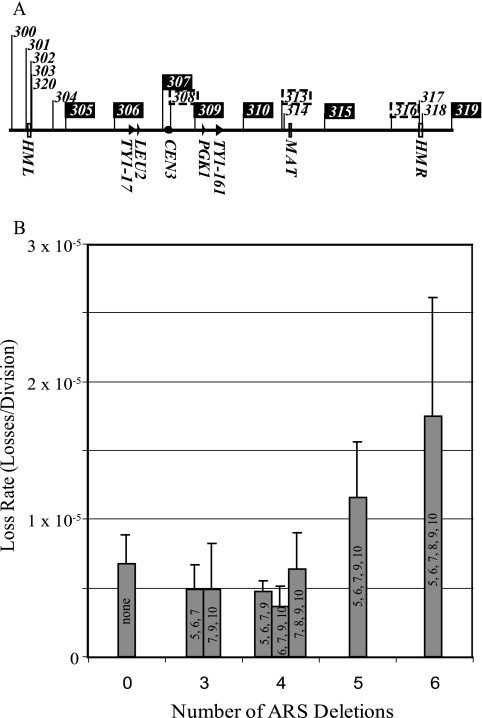
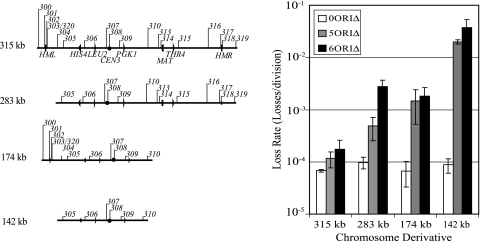
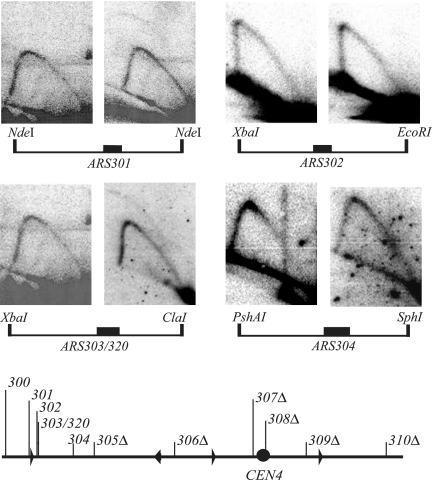
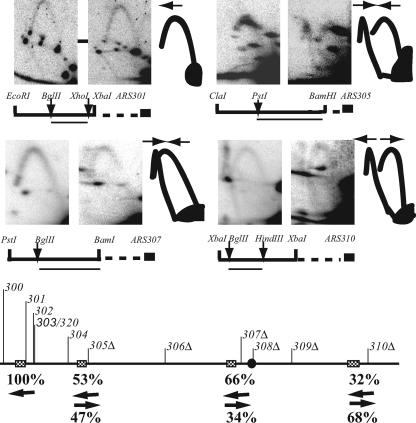
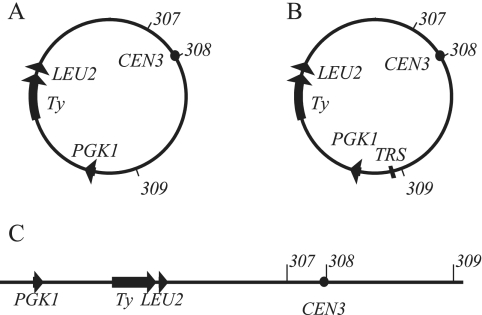
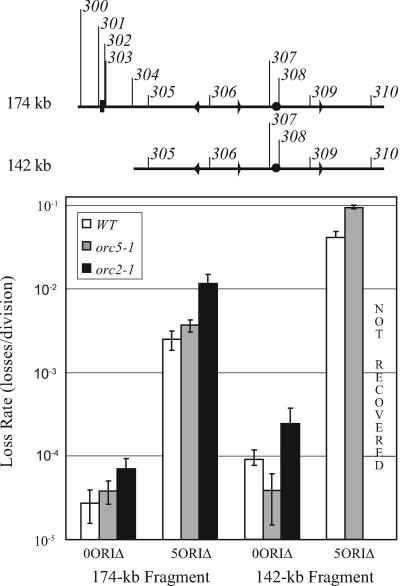
Replication origins in Saccharomyces cerevisiae are spaced at intervals of approximately 40 kb. However, both measurements of replication fork rate and studies of hypomorphic alleles of genes encoding replication initiation proteins suggest the question of whether replication origins are more closely spaced than should be required. We approached this question by systematically deleting replicators from chromosome III. The first significant increase in loss rate detected for the 315-kb full-length chromosome occurred only after all five efficient chromosomal replicators in the left two-thirds of the chromosome (ARS305, ARS306, ARS307, ARS309, and ARS310) had been deleted. The removal of the inefficient replicator ARS308 from this originless region caused little or no additional increase in loss rate. Chromosome fragmentations that removed the normally inactive replicators on the left end of the chromosome or the replicators distal to ARS310 on the right arm showed that both groups of replicators contribute significantly to the maintenance of the originless chromosome. Surprisingly, a 142-kb derivative of chromosome III, lacking all sequences that function as autonomously replicating sequence elements in plasmids, replicated and segregated properly 97% of the time. Both the replication initiation protein ORC and telomeres or a linear topology were required for the maintenance of chromosome fragments lacking replicators.
Figures
References
-
- Bell, S. P. 2002. The origin recognition complex: from simple origins to complex functions. Genes Dev. 16:659-672. - PubMed
-
- Bell, S. P., and A. Dutta. 2002. DNA replication in eukaryotic cells. Annu. Rev. Biochem. 71:333-374. - PubMed
-
- Boeke, J. D., J. Trueheart, G. Natsoulis, and G. R. Fink. 1987. 5-Fluoroorotic acid as a selective agent in yeast molecular genetics. Methods Enzymol. 154:164-175. - PubMed
-
- Brewer, B. J., and W. L. Fangman. 1988. A replication fork barrier at the 3′ end of yeast ribosomal RNA genes. Cell 55:637-643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases