

Ubiquitination-induced conformational change within the deiodinase dimer is a switch regulating enzyme activity
- PMID: 17452445
- PMCID: PMC1951476
- DOI: 10.1128/MCB.00283-07
Ubiquitination-induced conformational change within the deiodinase dimer is a switch regulating enzyme activity
Abstract
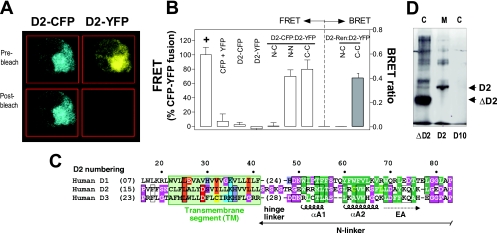
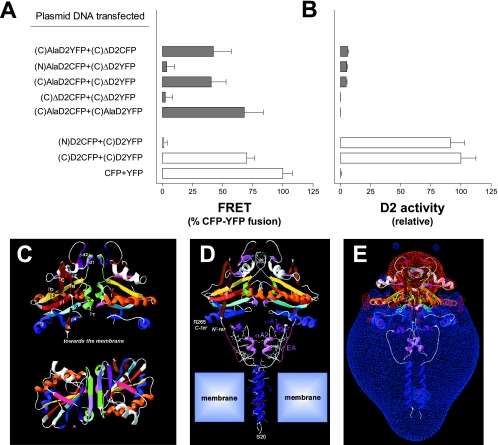
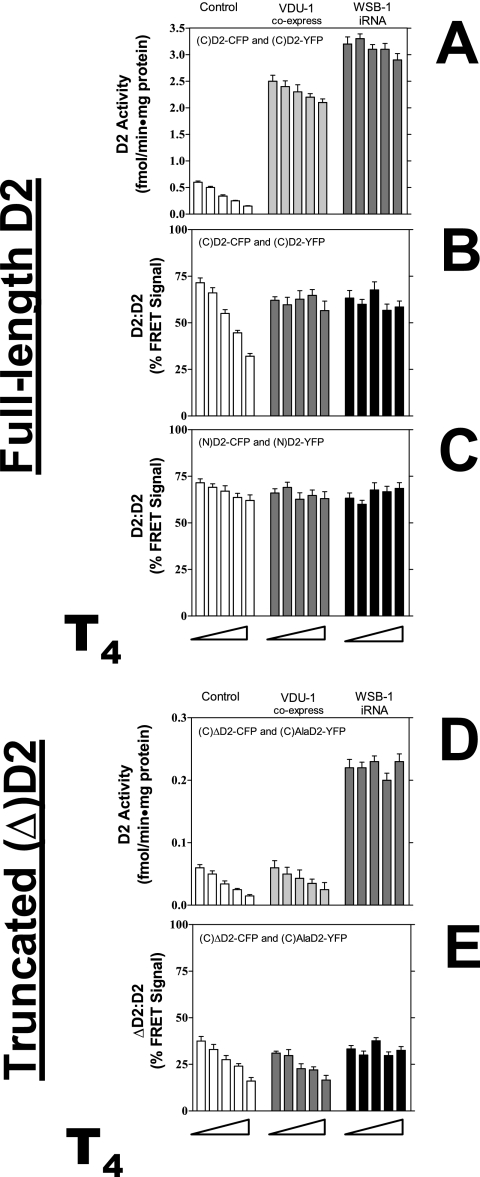
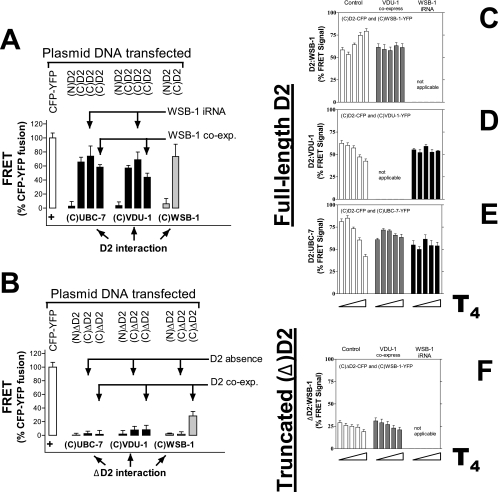
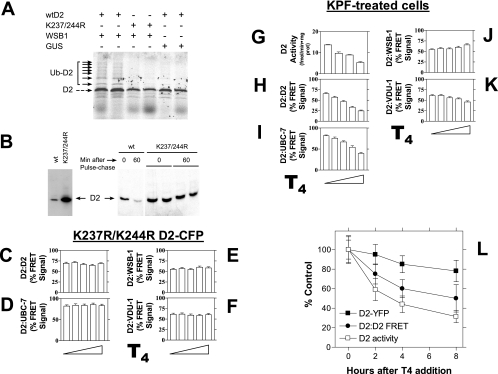
Ubiquitination is a critical posttranslational regulator of protein stability and/or subcellular localization. Here we show that ubiquitination can also regulate proteins by transiently inactivating enzymatic function through conformational change in a dimeric enzyme, which can be reversed upon deubiquitination. Our model system is the thyroid hormone-activating type 2 deiodinase (D2), an endoplasmic reticulum-resident type 1 integral membrane enzyme. D2 exists as a homodimer maintained by interacting surfaces at its transmembrane and globular cytosolic domains. The D2 dimer associates with the Hedgehog-inducible ubiquitin ligase WSB-1, the ubiquitin conjugase UBC-7, and VDU-1, a D2-specific deubiquitinase. Upon binding of T4, its natural substrate, D2 is ubiquitinated, which inactivates the enzyme by interfering with D2's globular interacting surfaces that are critical for dimerization and catalytic activity. This state of transient inactivity and change in dimer conformation persists until deubiquitination. The continuous association of D2 with this regulatory protein complex supports rapid cycles of deiodination, conjugation to ubiquitin, and enzyme reactivation by deubiquitination, allowing tight control of thyroid hormone action.
Figures
References
-
- Bahadur, R. P., P. Chakrabarti, F. Rodier, and J. Janin. 2004. A dissection of specific and non-specific protein-protein interfaces. J. Mol. Biol. 336:943-955. - PubMed
-
- Baqui, M., D. Botero, B. Gereben, C. Curcio, J. W. Harney, D. Salvatore, K. Sorimachi, P. R. Larsen, and A. C. Bianco. 2002. Human type 3 iodothyronine selenodeiodinase is located in the plasma membrane and undergoes rapid internalization to endosomes. J. Biol. Chem. 4:4. - PubMed
-
- Baqui, M. M., B. Gereben, J. W. Harney, P. R. Larsen, and A. C. Bianco. 2000. Distinct subcellular localization of transiently expressed types 1 and 2 iodothyronine deiodinases as determined by immunofluorescence confocal microscopy. Endocrinology 141:4309-4312. - PubMed
-
- Berry, M. J., A. L. Maia, J. D. Kieffer, J. W. Harney, and P. R. Larsen. 1992. Substitution of cysteine for selenocysteine in type I iodothyronine deiodinase reduces the catalytic efficiency of the protein but enhances its translation. Endocrinology 131:1848-1852. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials