

Unmasking activation of the zygotic genome using chromosomal deletions in the Drosophila embryo
- PMID: 17456005
- PMCID: PMC1854917
- DOI: 10.1371/journal.pbio.0050117
Unmasking activation of the zygotic genome using chromosomal deletions in the Drosophila embryo
Erratum in
- PLoS Biol. 2007 Aug;5(8):e213
- PLoS Biol. 2007 Jul;5(7):e195
Abstract
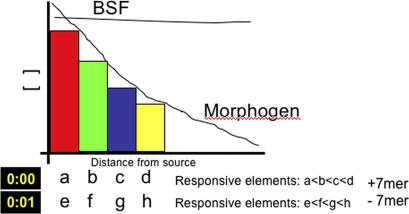
During the maternal-to-zygotic transition, a developing embryo integrates post-transcriptional regulation of maternal mRNAs with transcriptional activation of its own genome. By combining chromosomal ablation in Drosophila with microarray analysis, we characterized the basis of this integration. We show that the expression profile for at least one third of zygotically active genes is coupled to the concomitant degradation of the corresponding maternal mRNAs. The embryo uses transcription and degradation to generate localized patterns of expression, and zygotic transcription to degrade distinct classes of maternal transcripts. Although degradation does not appear to involve a simple regulatory code, the activation of the zygotic genome starts from intronless genes sharing a common cis-element. This cis-element interacts with a single protein, the Bicoid stability factor, and acts as a potent enhancer capable of timing the activity of an exogenous transactivator. We propose that this regulatory mode links morphogen gradients with temporal regulation during the maternal-to-zygotic transition.
Conflict of interest statement
Figures






Comment in
-
Novel technique shows when the fertilized egg's genome comes into its own.PLoS Biol. 2007 May;5(5):e131. doi: 10.1371/journal.pbio.0050131. Epub 2007 Apr 24. PLoS Biol. 2007. PMID: 20076672 Free PMC article. No abstract available.
Similar articles
-
The conserved regulatory basis of mRNA contributions to the early Drosophila embryo differs between the maternal and zygotic genomes.PLoS Genet. 2020 Mar 30;16(3):e1008645. doi: 10.1371/journal.pgen.1008645. eCollection 2020 Mar. PLoS Genet. 2020. PMID: 32226006 Free PMC article.
-
The zinc-finger protein Zelda is a key activator of the early zygotic genome in Drosophila.Nature. 2008 Nov 20;456(7220):400-3. doi: 10.1038/nature07388. Epub 2008 Oct 19. Nature. 2008. PMID: 18931655 Free PMC article.
-
Transcriptional and post-transcriptional regulation of maternal and zygotic cytoskeletal tropomyosin mRNA during Drosophila development correlates with specific morphogenic events.Dev Biol. 1994 Oct;165(2):639-53. doi: 10.1006/dbio.1994.1282. Dev Biol. 1994. PMID: 7958428
-
Regulatory principles governing the maternal-to-zygotic transition: insights from Drosophila melanogaster.Open Biol. 2018 Dec;8(12):180183. doi: 10.1098/rsob.180183. Open Biol. 2018. PMID: 30977698 Free PMC article. Review.
-
Regulation and Function of Maternal Gene Products During the Maternal-to-Zygotic Transition in Drosophila.Curr Top Dev Biol. 2015;113:43-84. doi: 10.1016/bs.ctdb.2015.06.007. Epub 2015 Aug 14. Curr Top Dev Biol. 2015. PMID: 26358870 Review.
Cited by
-
Balancing sex chromosome expression and satisfying the sexes.Fly (Austin). 2012 Jan-Mar;6(1):26-9. doi: 10.4161/fly.18822. Epub 2012 Jan 1. Fly (Austin). 2012. PMID: 22388008 Free PMC article.
-
Mechanisms regulating zygotic genome activation.Nat Rev Genet. 2019 Apr;20(4):221-234. doi: 10.1038/s41576-018-0087-x. Nat Rev Genet. 2019. PMID: 30573849 Free PMC article. Review.
-
REDfly: the transcriptional regulatory element database for Drosophila.Nucleic Acids Res. 2019 Jan 8;47(D1):D828-D834. doi: 10.1093/nar/gky957. Nucleic Acids Res. 2019. PMID: 30329093 Free PMC article.
-
dFMRP and Caprin, translational regulators of synaptic plasticity, control the cell cycle at the Drosophila mid-blastula transition.Development. 2010 Dec;137(24):4201-9. doi: 10.1242/dev.055046. Epub 2010 Nov 10. Development. 2010. PMID: 21068064 Free PMC article.
-
Smaug destroys a huge treasure.Genome Biol. 2014 Jan 27;15(1):101. doi: 10.1186/gb4156. Genome Biol. 2014. PMID: 24468051 Free PMC article.
References
-
- Newport J, Kirschner M. A major developmental transition in early Xenopus embryos: II. Control of the onset of transcription. Cell. 1982;30:687–696. - PubMed
-
- Hamatani T, Carter MG, Sharov AA, Ko MS. Dynamics of global gene expression changes during mouse preimplantation development. Dev Cell. 2004;6:117–131. - PubMed
-
- Mathavan S, Lee SG, Mak A, Miller LD, Murthy KR, et al. Transcriptome analysis of zebrafish embryogenesis using microarrays. PLoS Genet. 2005;1:260–276. doi: 10.1371/journal.pgen.0010029. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
