

Viral alteration of cellular translational machinery increases defective ribosomal products
- PMID: 17459927
- PMCID: PMC1933321
- DOI: 10.1128/JVI.00137-07
Viral alteration of cellular translational machinery increases defective ribosomal products
Abstract
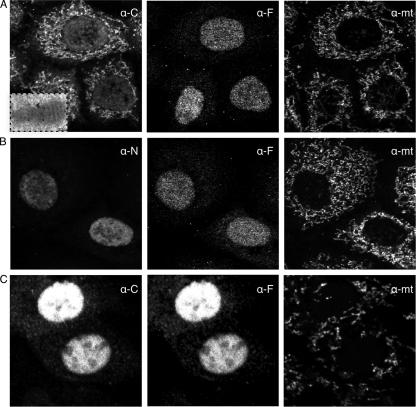
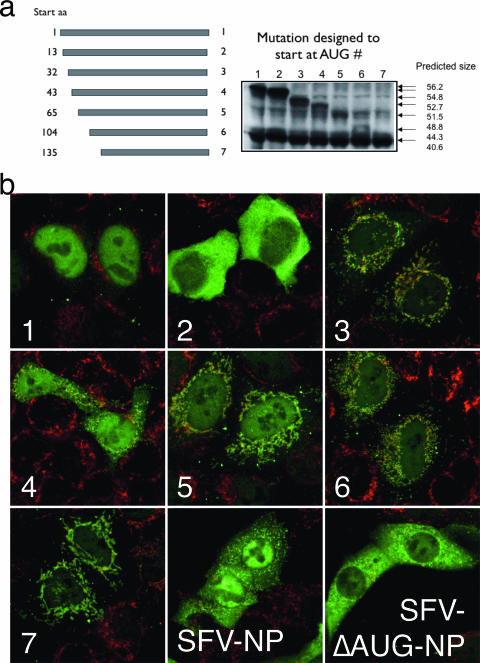
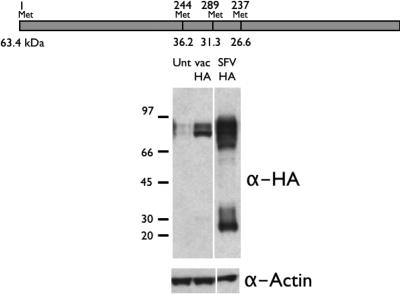
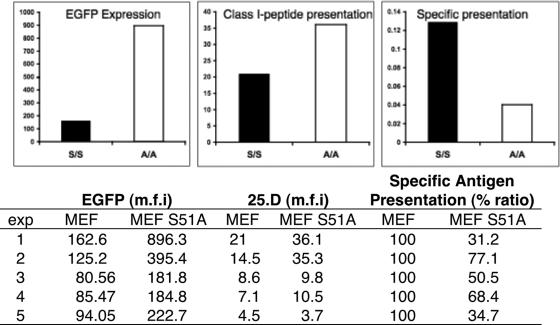
Here we show that cells expressing genes inserted into Semliki Forest virus (SFV) vectors generate a large fraction of defective ribosomal products (DRiPs) due to frequent initiation on downstream Met residues. In monopolizing the host cell translational machinery, SFV reduces levels of translation eukaryotic initiation factor 4E (eIF4E), diminishes phosphorylation of ribosome subunit S6, and phosphorylates translation initiation factor eIF2alpha. We show that the last event is required for SFV mistranslation of inserted genes. Downstream initiation is suppressed by fusing inserted genes with the open reading frame encoding the SFV capsid, demonstrating that one function of the capsid element is to enable ribosomes to initiate translation in the proper location. These results show that in modifying translation, viral vectors can unpredictably increase the generation of truncated polypeptides and thereby the DRiP fraction of inserted gene products, which can potentially affect their yield, therapeutic efficacy, and immunogenicity.
Figures



References
-
- Berglund, P., M. N. Fleeton, C. Smerdou, and P. Liljestrom. 1999. Immunization with recombinant Semliki Forest virus induces protection against influenza challenge in mice. Vaccine 17:497-507. - PubMed
-
- Berglund, P., M. Sjoberg, H. Garoff, G. J. Atkins, B. J. Sheahan, and P. Liljestrom. 1993. Semliki Forest virus expression system: production of conditionally infectious recombinant particles. Bio/Technology 11:916-920. - PubMed
-
- Berglund, P., C. Smerdou, M. N. Fleeton, I. Tubulekas, and P. Liljestrom. 1998. Enhancing immune responses using suicidal DNA vaccines. Nat. Biotechnol. 16:562-565. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
