

Localization of HCN1 channels to presynaptic compartments: novel plasticity that may contribute to hippocampal maturation
- PMID: 17460082
- PMCID: PMC3086816
- DOI: 10.1523/JNEUROSCI.4699-06.2007
Localization of HCN1 channels to presynaptic compartments: novel plasticity that may contribute to hippocampal maturation
Abstract
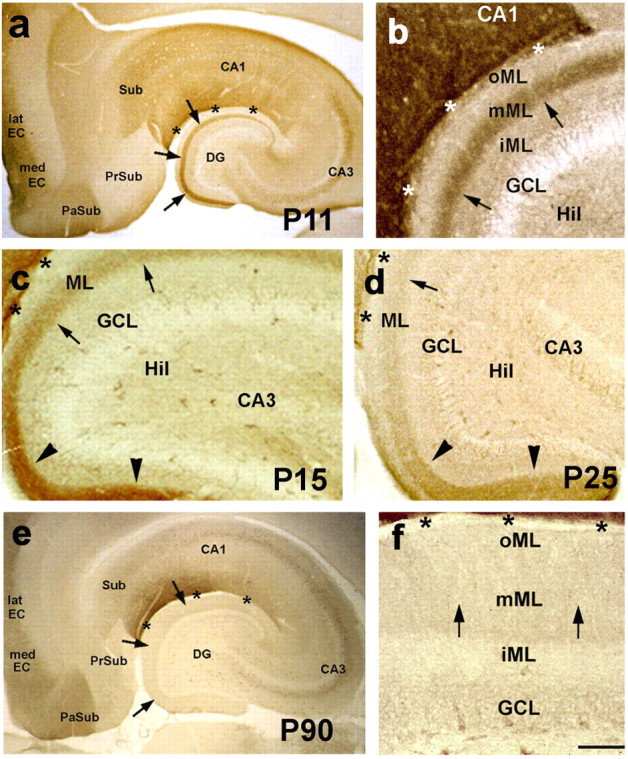
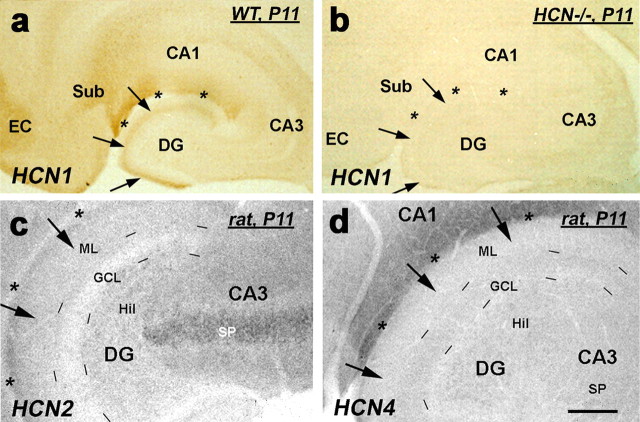
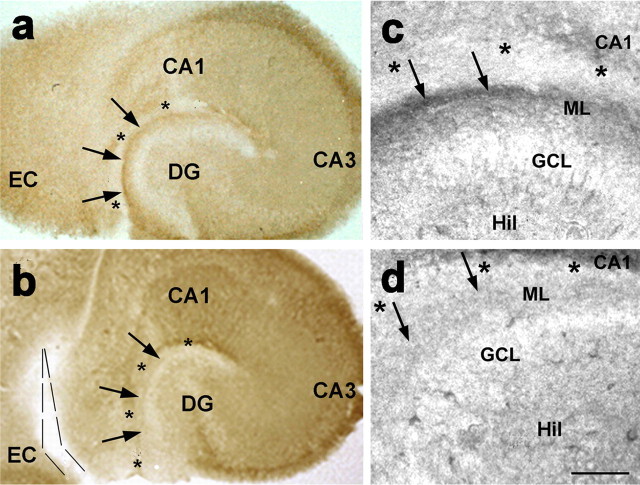
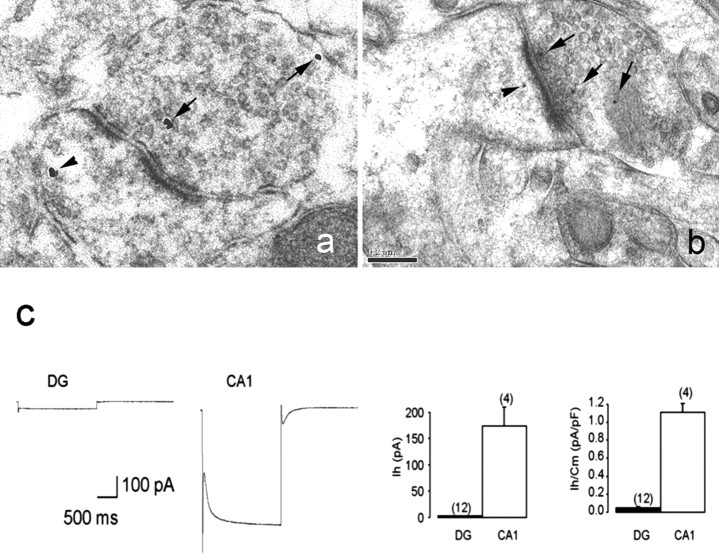
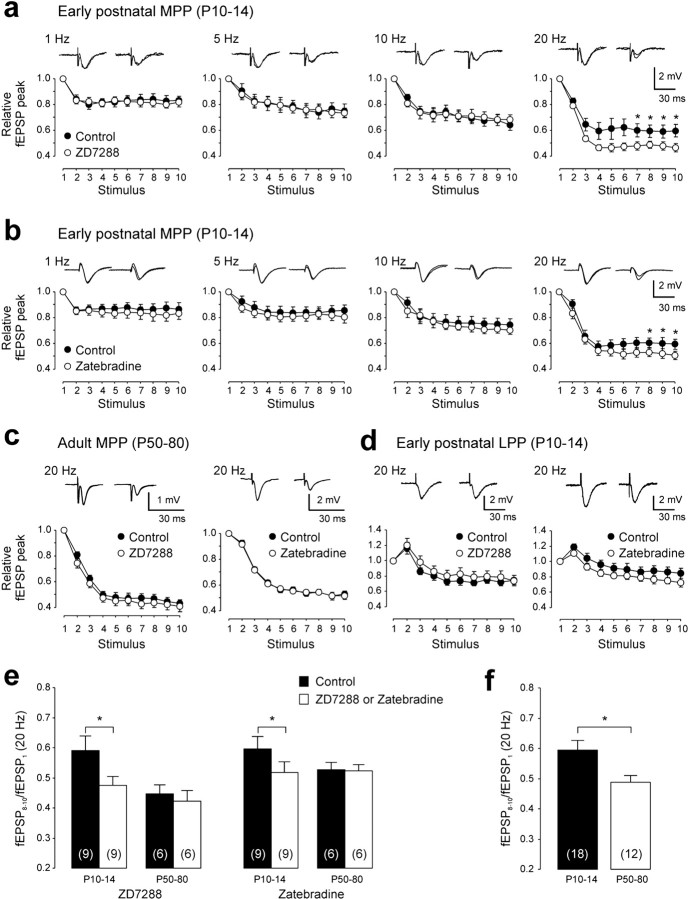
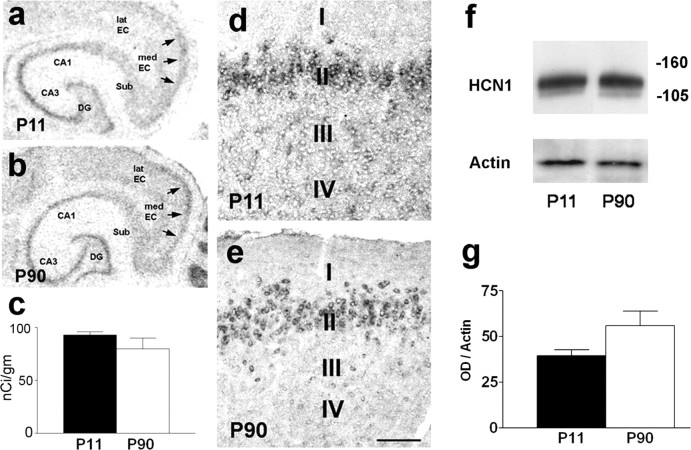
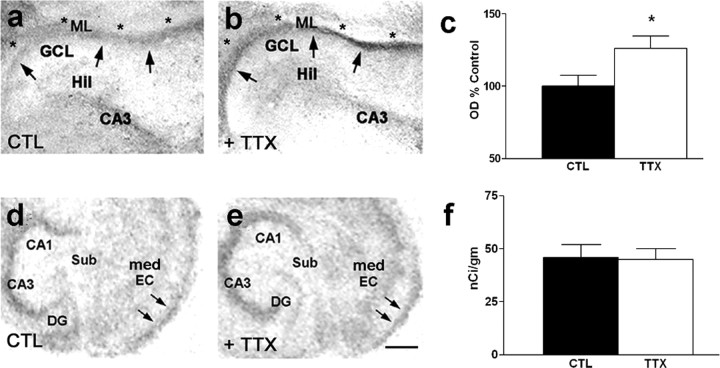
Increasing evidence supports roles for the current mediated by hyperpolarization-activated cyclic nucleotide-gated (HCN) channels, I(h), in hippocampal maturation and specifically in the evolving changes of intrinsic properties as well as network responses of hippocampal neurons. Here, we describe a novel developmental plasticity of HCN channel expression in axonal and presynaptic compartments: HCN1 channels were localized to axon terminals of the perforant path (the major hippocampal afferent pathway) of immature rats, where they modulated synaptic efficacy. However, presynaptic expression and functions of the channels disappeared with maturation. This was a result of altered channel transport to the axons, because HCN1 mRNA and protein levels in entorhinal cortex neurons, where the perforant path axons originate, were stable through adulthood. Blocking action potential firing in vitro increased presynaptic expression of HCN1 channels in the perforant path, suggesting that network activity contributed to regulating this expression. These findings support a novel developmentally regulated axonal transport of functional ion channels and suggest a role for HCN1 channel-mediated presynaptic I(h) in hippocampal maturation.
Figures
References
-
- Amaral DG, Witter MP. The hippocampal formation. In: Paxinos G, editor. The rat nervous system. Ed 2. New York: Academic; 1995. pp. 443–494.
-
- Beaumont V, Zucker RS. Enhancement of synaptic transmission by cyclic AMP modulation of presynaptic Ih channels. Nat Neurosci. 2000;3:133–141. - PubMed
-
- Ben-Ari Y. Excitatory actions of GABA during development: the nature of the nurture. Nat Rev Neurosci. 2002;3:728–739. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases