

Changes in the structural complexity of the aged brain
- PMID: 17465981
- PMCID: PMC2441530
- DOI: 10.1111/j.1474-9726.2007.00289.x
Changes in the structural complexity of the aged brain
Abstract
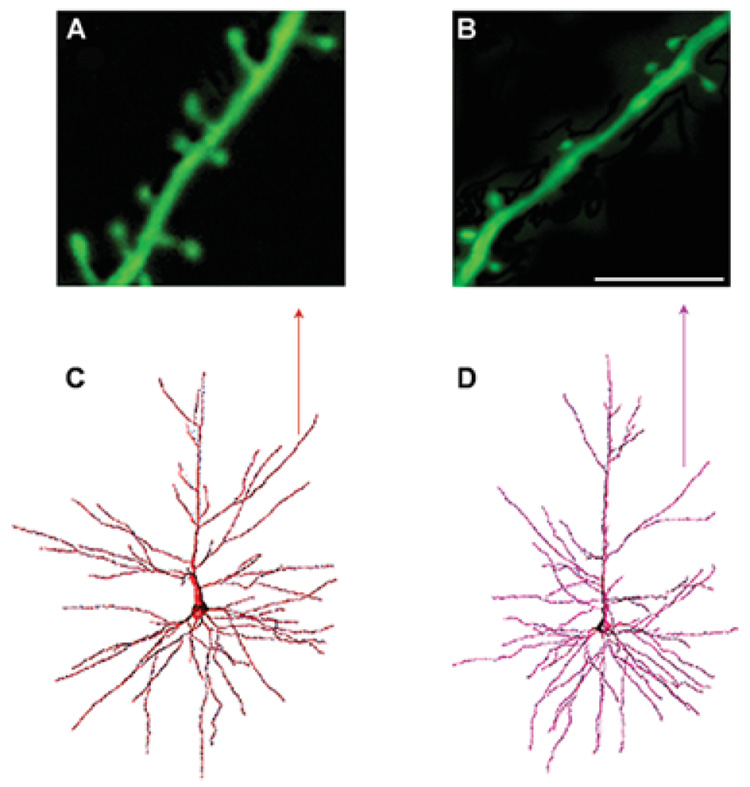
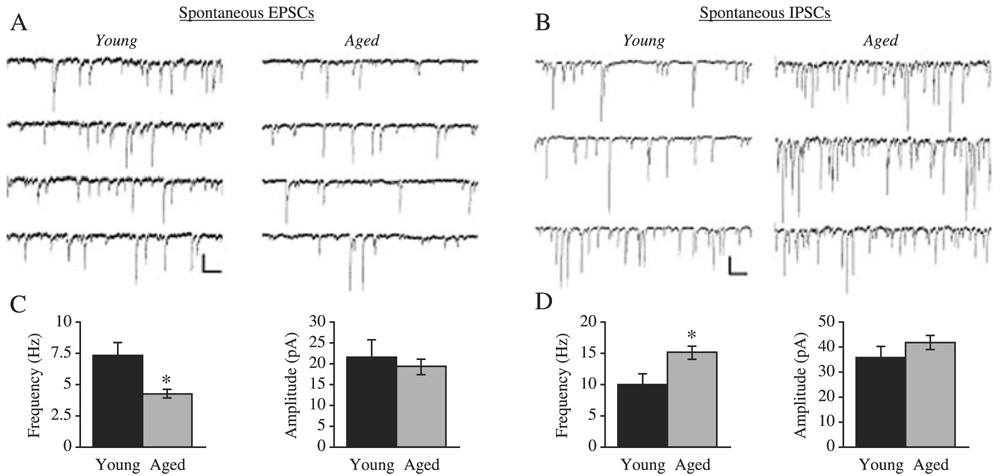
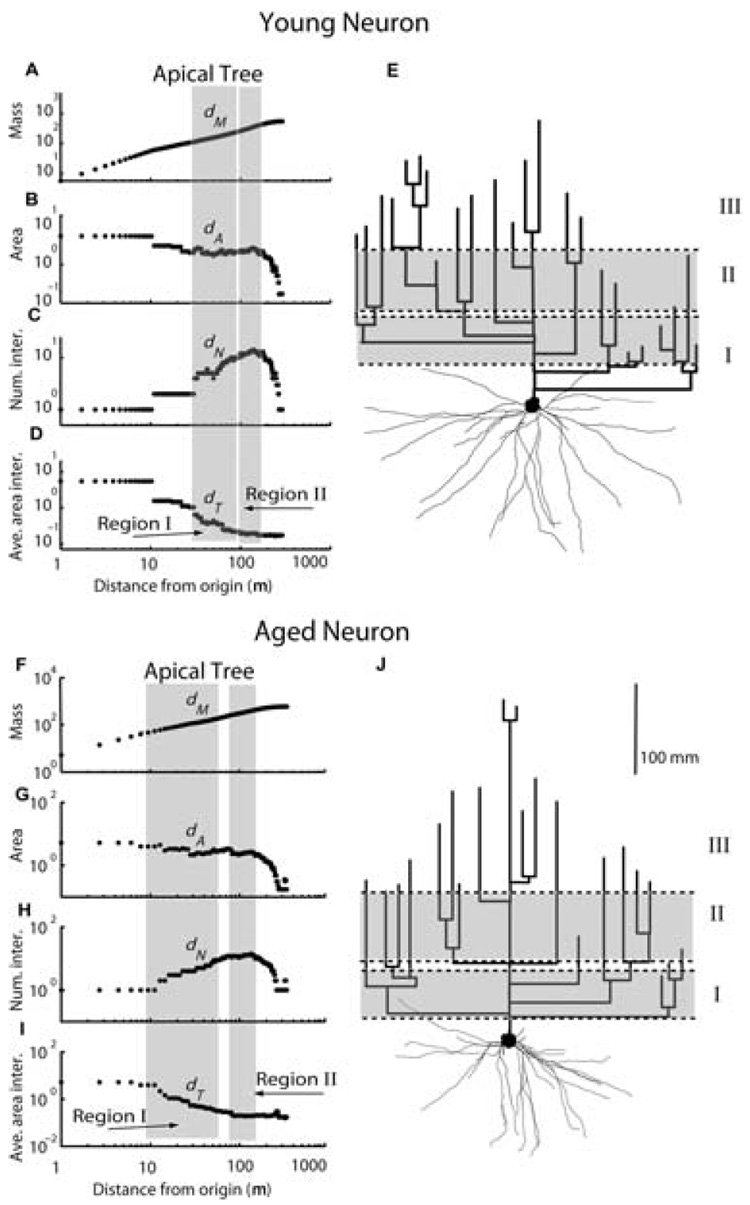
Structural changes of neurons in the brain during aging are complex and not well understood. Neurons have significant homeostatic control of essential brain functions, including synaptic excitability, gene expression, and metabolic regulation. Any deviations from the norm can have severe consequences as seen in aging and injury. In this review, we present some of the structural adaptations that neurons undergo throughout normal and pathological aging and discuss their effects on electrophysiological properties and cognition. During aging, it is evident that neurons undergo morphological changes such as a reduction in the complexity of dendrite arborization and dendritic length. Spine numbers are also decreased, and because spines are the major sites for excitatory synapses, changes in their numbers could reflect a change in synaptic densities. This idea has been supported by studies that demonstrate a decrease in the overall frequency of spontaneous glutamate receptor-mediated excitatory responses, as well as a decrease in the levels of alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid and N-methyl-d-aspartate receptor expression. Other properties such as gamma-aminobutyric acid A receptor-mediated inhibitory responses and action potential firing rates are both significantly increased with age. These findings suggest that age-related neuronal dysfunction, which must underlie observed decline in cognitive function, probably involves a host of other subtle changes within the cortex that could include alterations in receptors, loss of dendrites, and spines and myelin dystrophy, as well as the alterations in synaptic transmission. Together these multiple alterations in the brain may constitute the substrate for age-related loss of cognitive function.
Figures
References
-
- Alpar A, Ueberham U, Bruckner MK, Arendt T, Gartner U. The expression of wild-type human amyloid precursor protein affects the dendritic phenotype of neocortical pyramidal neurons in transgenic mice. Int. J. Dev. Neurosci. 2006a;24:133–140. - PubMed
-
- Alpar A, Ueberham U, Bruckner MK, Seeger G, Arendt T, Gartner U. Different dendrite and dendritic spine alterations in basal and apical arbors in mutant human amyloid precursor protein transgenic mice. Brain Res. 2006b;1099:189–198. - PubMed
-
- Andorfer C, Kress Y, Espinoza M, de Silva R, Tucker KL, Barde YA, Duff K, Davies P. Hyperphosphorylation and aggregation of tau in mice expressing normal human tau isoforms. J. Neurochem. 2003;86:582–590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
