

The COMATOSE ATP-binding cassette transporter is required for full fertility in Arabidopsis
- PMID: 17468211
- PMCID: PMC1914130
- DOI: 10.1104/pp.107.099903
The COMATOSE ATP-binding cassette transporter is required for full fertility in Arabidopsis
Abstract
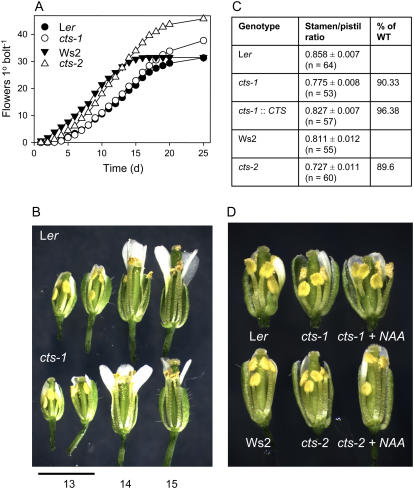
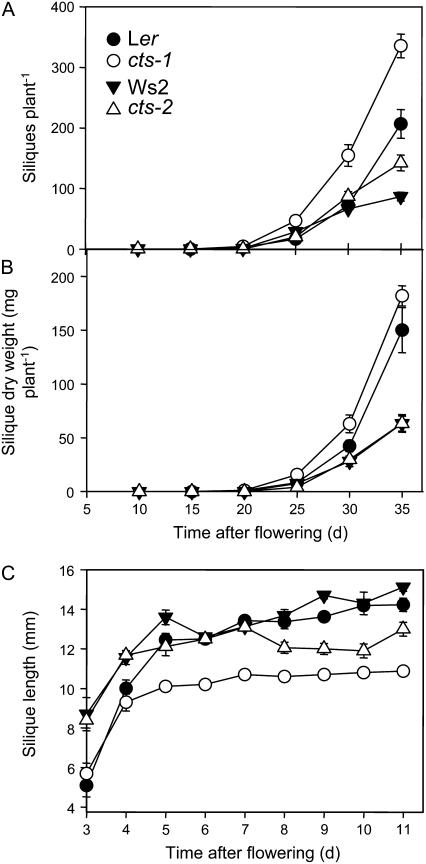
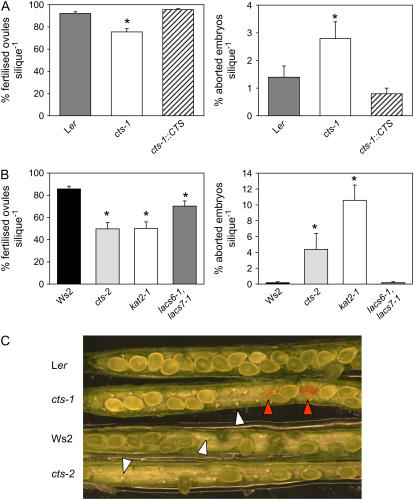
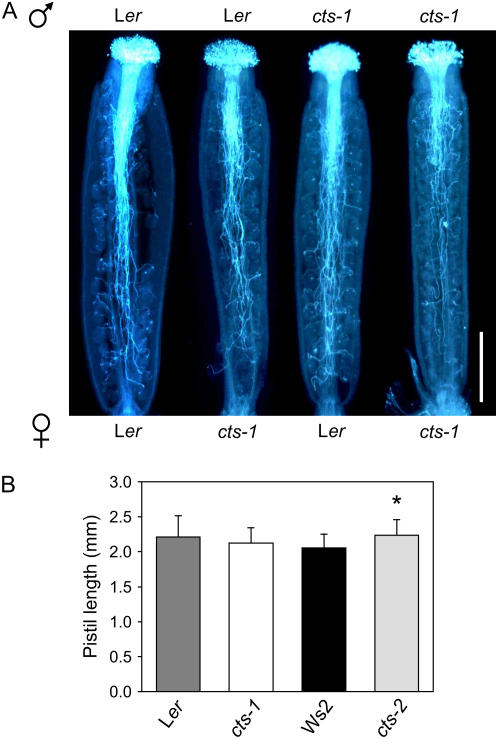
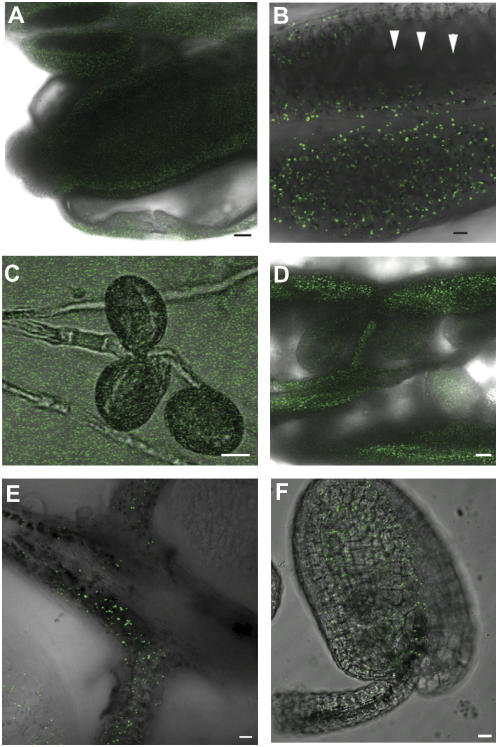
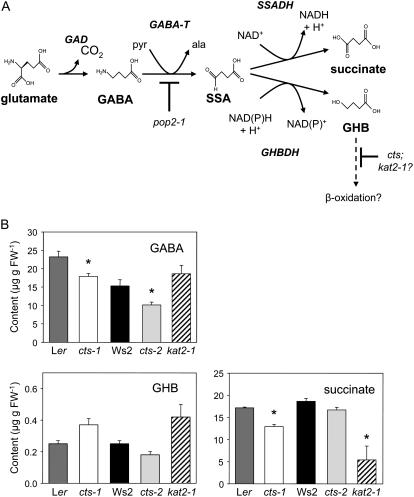
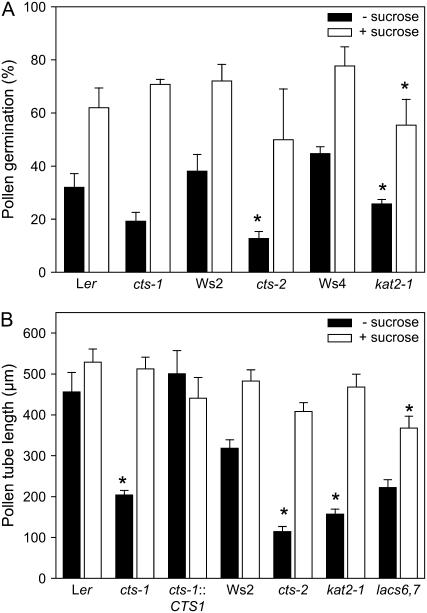
COMATOSE (CTS) encodes a peroxisomal ATP-binding cassette transporter required not only for beta-oxidation of storage lipids during germination and establishment, but also for biosynthesis of jasmonic acid and conversion of indole butyric acid to indole acetic acid. cts mutants exhibited reduced fertilization, which was rescued by genetic complementation, but not by exogenous application of jasmonic acid or indole acetic acid. Reduced fertilization was also observed in thiolase (kat2-1) and peroxisomal acyl-Coenzyme A synthetase mutants (lacs6-1,lacs7-1), indicating a general role for beta-oxidation in fertility. Genetic analysis revealed reduced male transmission of cts alleles and both cts pollen germination and tube growth in vitro were impaired in the absence of an exogenous carbon source. Aniline blue staining of pollinated pistils demonstrated that pollen tube growth was affected only when both parents bore the cts mutation, indicating that expression of CTS in either male or female tissues was sufficient to support pollen tube growth in vivo. Accordingly, abundant peroxisomes were detected in a range of maternal tissues. Although gamma-aminobutyric acid levels were reduced in flowers of cts mutants, they were unchanged in kat2-1, suggesting that alterations in gamma-aminobutyric acid catabolism do not contribute to the reduced fertility phenotype through altered pollen tube targeting. Taken together, our data support an important role for beta-oxidation in fertility in Arabidopsis (Arabidopsis thaliana) and suggest that this pathway could play a role in the mobilization of lipids in both pollen and female tissues.
Figures
Similar articles
-
Analysis of the role of COMATOSE and peroxisomal beta-oxidation in the determination of germination potential in Arabidopsis.J Exp Bot. 2006;57(11):2805-14. doi: 10.1093/jxb/erl045. Epub 2006 Jul 14. J Exp Bot. 2006. PMID: 16844736
-
Jasmonic acid levels are reduced in COMATOSE ATP-binding cassette transporter mutants. Implications for transport of jasmonate precursors into peroxisomes.Plant Physiol. 2005 Mar;137(3):835-40. doi: 10.1104/pp.105.059352. Plant Physiol. 2005. PMID: 15761209 Free PMC article. No abstract available.
-
Sucrose rescues seedling establishment but not germination of Arabidopsis mutants disrupted in peroxisomal fatty acid catabolism.Plant J. 2005 Sep;43(6):861-72. doi: 10.1111/j.1365-313X.2005.02498.x. Plant J. 2005. PMID: 16146525
-
Mutations in the Arabidopsis peroxisomal ABC transporter COMATOSE allow differentiation between multiple functions in planta: insights from an allelic series.Mol Biol Cell. 2009 Jan;20(1):530-43. doi: 10.1091/mbc.e08-07-0745. Epub 2008 Nov 19. Mol Biol Cell. 2009. PMID: 19019987 Free PMC article.
-
Peroxisomal Acyl-CoA synthetase activity is essential for seedling development in Arabidopsis thaliana.Plant Cell. 2004 Feb;16(2):394-405. doi: 10.1105/tpc.019646. Epub 2004 Jan 23. Plant Cell. 2004. PMID: 14742880 Free PMC article.
Cited by
-
Jasmonate signaling in plant development and defense response to multiple (a)biotic stresses.Plant Cell Rep. 2013 Jul;32(7):1085-98. doi: 10.1007/s00299-013-1441-2. Epub 2013 Apr 13. Plant Cell Rep. 2013. PMID: 23584548 Review.
-
Seed storage oil mobilization is important but not essential for germination or seedling establishment in Arabidopsis.Plant Physiol. 2011 Oct;157(2):866-75. doi: 10.1104/pp.111.181784. Epub 2011 Aug 8. Plant Physiol. 2011. PMID: 21825108 Free PMC article.
-
Defects in Peroxisomal 6-Phosphogluconate Dehydrogenase Isoform PGD2 Prevent Gametophytic Interaction in Arabidopsis thaliana.Plant Physiol. 2016 May;171(1):192-205. doi: 10.1104/pp.15.01301. Epub 2016 Mar 3. Plant Physiol. 2016. PMID: 26941195 Free PMC article.
-
Functional characterization of a higher plant sphingolipid Delta4-desaturase: defining the role of sphingosine and sphingosine-1-phosphate in Arabidopsis.Plant Physiol. 2009 Jan;149(1):487-98. doi: 10.1104/pp.108.129411. Epub 2008 Oct 31. Plant Physiol. 2009. PMID: 18978071 Free PMC article.
-
Importance of organellar proteins, protein translocation and vesicle transport routes for pollen development and function.Plant Reprod. 2016 Jun;29(1-2):53-65. doi: 10.1007/s00497-016-0274-x. Epub 2016 Feb 13. Plant Reprod. 2016. PMID: 26874709 Review.
References
-
- Afitlhile MM, Fukushige H, Nishimura M, Hildebrand DF (2005) A defect in glyoxysomal fatty acid beta-oxidation reduces jasmonic acid accumulation in Arabidopsis. Plant Physiol Biochem 43 603–609 - PubMed
-
- Alexander MP (1969) Differential staining of aborted and nonaborted pollen. Stain Technol 44 117–122 - PubMed
-
- Aloni R, Aloni E, Langhans M, Ullrich CI (2006) Role of auxin in regulating Arabidopsis flower development. Planta 223 315–328 - PubMed
-
- Baker A, Graham IA, Holdsworth M, Smith SJ, Theodoulou FL (2006) Chewing the fat: β-oxidation in signalling and development. Trends Plant Sci 11 124–132 - PubMed
-
- Baker HG, Baker I (1979) Starch in angiosperm pollen grains and its evolutionary significance. Am J Bot 66 591–600
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
