

Plant mitochondrial recombination surveillance requires unusual RecA and MutS homologs
- PMID: 17468263
- PMCID: PMC1913765
- DOI: 10.1105/tpc.106.048355
Plant mitochondrial recombination surveillance requires unusual RecA and MutS homologs
Abstract
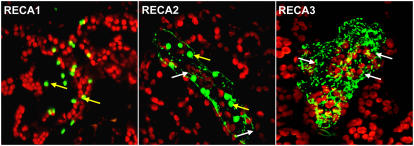
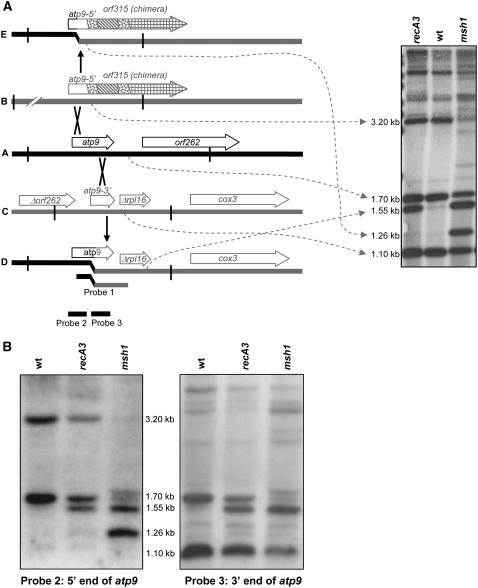
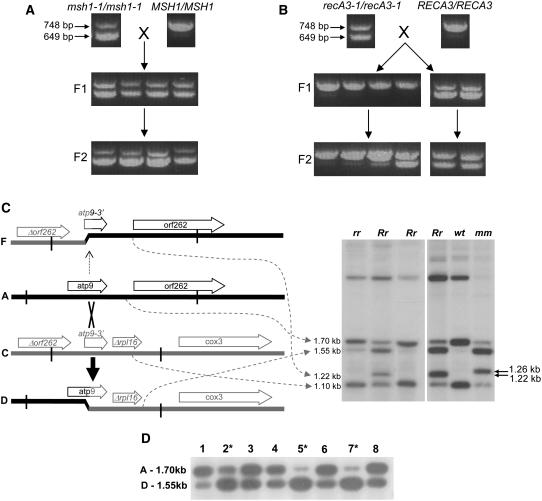
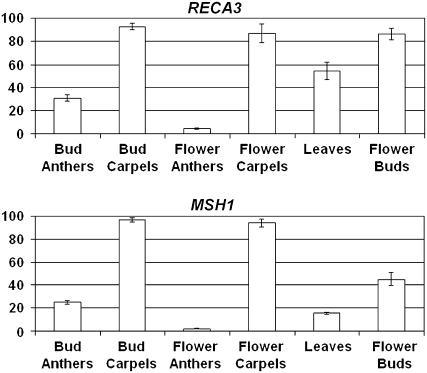
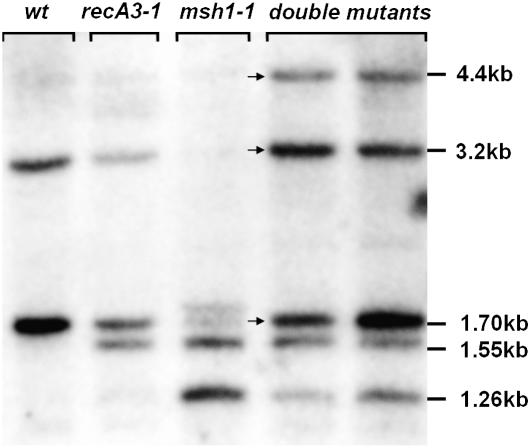
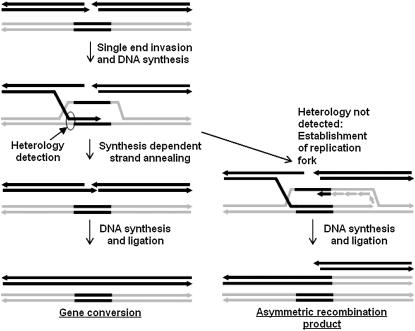
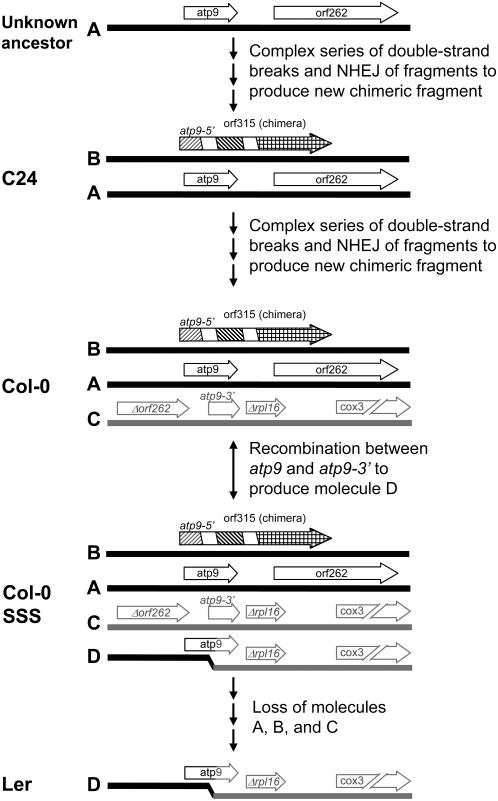
For >20 years, the enigmatic behavior of plant mitochondrial genomes has been well described but not well understood. Chimeric genes appear, and occasionally are differentially replicated or expressed, with significant effects on plant phenotype, most notably on male fertility, yet the mechanisms of DNA replication, chimera formation, and recombination have remained elusive. Using mutations in two important genes of mitochondrial DNA metabolism, we have observed reproducible asymmetric recombination events occurring at specific locations in the mitochondrial genome. Based on these experiments and existing models of double-strand break repair, we propose a model for plant mitochondrial DNA replication, chimeric gene formation, and the illegitimate recombination events that lead to stoichiometric changes. We also address the physiological and developmental effects of aberrant events in mitochondrial genome maintenance, showing that mitochondrial genome rearrangements, when controlled, influence plant reproduction, but when uncontrolled, lead to aberrant growth phenotypes and dramatic reduction of the cell cycle.
Figures


Similar articles
-
Mitochondrial genome dynamics in plants and animals: convergent gene fusions of a MutS homologue.J Mol Evol. 2006 Aug;63(2):165-73. doi: 10.1007/s00239-005-0226-9. Epub 2006 Jul 7. J Mol Evol. 2006. PMID: 16830098
-
Double-strand break repair processes drive evolution of the mitochondrial genome in Arabidopsis.BMC Biol. 2011 Sep 27;9:64. doi: 10.1186/1741-7007-9-64. BMC Biol. 2011. PMID: 21951689 Free PMC article.
-
MSH1 is required for maintenance of the low mutation rates in plant mitochondrial and plastid genomes.Proc Natl Acad Sci U S A. 2020 Jul 14;117(28):16448-16455. doi: 10.1073/pnas.2001998117. Epub 2020 Jun 29. Proc Natl Acad Sci U S A. 2020. PMID: 32601224 Free PMC article.
-
The bacterial RecA protein and the recombinational DNA repair of stalled replication forks.Annu Rev Biochem. 2002;71:71-100. doi: 10.1146/annurev.biochem.71.083101.133940. Epub 2001 Nov 9. Annu Rev Biochem. 2002. PMID: 12045091 Review.
-
The plant mitochondrial genome: dynamics and maintenance.Biochimie. 2014 May;100:107-20. doi: 10.1016/j.biochi.2013.09.016. Epub 2013 Sep 26. Biochimie. 2014. PMID: 24075874 Review.
Cited by
-
Long-read sequencing characterizes mitochondrial and plastid genome variants in Arabidopsis msh1 mutants.Plant J. 2022 Nov;112(3):738-755. doi: 10.1111/tpj.15976. Epub 2022 Sep 21. Plant J. 2022. PMID: 36097957 Free PMC article.
-
Mosaic origins of a complex chimeric mitochondrial gene in Silene vulgaris.PLoS One. 2012;7(2):e30401. doi: 10.1371/journal.pone.0030401. Epub 2012 Feb 27. PLoS One. 2012. PMID: 22383961 Free PMC article.
-
Mitotypes Based on Structural Variation of Mitochondrial Genomes Imply Relationships With Morphological Phenotypes and Cytoplasmic Male Sterility in Peppers.Front Plant Sci. 2019 Oct 24;10:1343. doi: 10.3389/fpls.2019.01343. eCollection 2019. Front Plant Sci. 2019. PMID: 31708952 Free PMC article.
-
Genetic Diversity, Heteroplasmy, and Recombination in Mitochondrial Genomes of Daphnia pulex, Daphnia pulicaria, and Daphnia obtusa.Mol Biol Evol. 2022 Apr 11;39(4):msac059. doi: 10.1093/molbev/msac059. Mol Biol Evol. 2022. PMID: 35325186 Free PMC article.
-
Characterization of a novel thermosensitive restorer of fertility for cytoplasmic male sterility in maize.Genetics. 2009 May;182(1):91-103. doi: 10.1534/genetics.108.099895. Epub 2009 Mar 2. Genetics. 2009. PMID: 19255365 Free PMC article.
References
-
- Abdelnoor, R., Christensen, A., Mohammed, S., Munoz-Castillo, B., Moriyama, H., and Mackenzie, S. (2006). Mitochondrial genome dynamics in plants and animals: Convergent gene fusions of a MutS homologue. J. Mol. Evol. 63 165–173. - PubMed
-
- Adams, K., and Palmer, J. (2003). Evolution of mitochondrial gene content: Gene loss and transfer to the nucleus. Mol. Phylogenet. Evol. 29 380–395. - PubMed
-
- Aguilera, A. (2001). Double-strand break repair: Are Rad51/RecA–DNA joints barriers to DNA replication? Trends Genet. 17 318–321. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
